



Акустические особенности насекомых имеют важнейшее значение при размножении, благодаря чему обеспечивается встреча особей противоположного пола и репродуктивная изоляция близких симпатрических видов [1]. Неослабевающий интерес к звуковой коммуникации комаров объясняется как желанием понять наиболее сложные формы их жизнедеятельности, так и необходимостью управлять поведением и контролировать численность переносчиков опасных трансмиссивных заболеваний.
Малярийные комары рода Anopheles (Diptera, Culicidae) являются объектами постоянного повышенного внимания со стороны биологов и медицинских служб ввиду эпидемиологического значения этих насекомых. В то же время представители данной группы являются прекрасными модельными объектами для генетического и экологического анализа [2, 6].
Цитогенетические исследования малярийных комаров Палеарктики позволили выявить ряд видов-двойников, объединенных в комплекс «Anophelesmaculipennis», для некоторых из которых характерен внутривидовой инверсионно-хромосомный полиморфизм, имеющий адаптивное значение [9]. Несмотря на пристальное внимание к малярийным комарам со стороны специалистов, механизмы звуковой коммуникации этой группы двукрылых изучены недостаточно.
Целью настоящей работы являлось изучение акустических механизмов идентификации и выбора полового партнера у видов-двойников малярийных комаров комплекса «Anophelesmaculipennis».
Материал и методы
Материалом для данной работы послужили комары четырех видов-двойников: An. messeaeFalleroni, An. beklemisheviStegniyandKabanova, An. maculipennisMeigen и An. atroparvusvanThiel. Имаго An. atroparvus использованы из лабораторной линии; имаго остальных видов выведены из потомств от гоноактивных самок, выборки которых были взяты в природных популяциях – An. messeae иAn. beklemishevi в Томской области, An. maculipennis в Белоруссии. Среди использованных потомств An. beklemishevi, An. atroparvus и An. maculipennis оказались цитогенетически мономорфными, а An. messeae – инверсионно полиморфным. Поэтому в последнем случае цитогенетически определяли кариотипы особей из каждого потомства. У выращенных имаго всех четырех видов снимались акустические характеристики. Для этого комаров приклеивали дорсальной стороной груди к препаровальной игле (рис. 1) и помещали в звуковую камеру с микрофоном (рис. 2). Для регистрации акустических сигналов использовался компьютер и соответствующее программное обеспечение.

Рисунок 1.Фиксация комара перед микрофоном

Рисунок 2.Звуковая камера с микрофоном
Содержание личинок всех видов осуществлялось в одних и тех же условиях и на одной и той же диете. Статистическая обработка результатов исследований проводилась средствами MicrosoftOfficeExcel с анализом «выбросов», проверки дисперсионной однородности и соответствия выборок нормальному распределению.
Результаты и обсуждение
В различных регионах Палеарктики установлена симпатрия представителей малярийных комаров в разнообразных сочетаниях [3, 6]. В связи с этим в процессе эволюции у комаров должны были сформироваться четкие биологические докопуляционные механизмы, обеспечивающие встречу и распознавание особей противоположного пола своего вида для скрещивания. Учитывая то, что скрещивание комаров происходит в полете и не редко в темное время суток [2], первостепенное значение в видовой идентификации, очевидно, имеют акустические сигналы, издаваемые машущими крыльями.
Для комаров Aedesaegypti [8] и Culexquinquefasciatus [10] было показано, что в их звуковом спектре при идентификации и выборе полового партнера определяющее значение имеет не только величина основного тона, но и кратные ей по частоте гармоники. Амплитуды информативных сигналов и низкочастотные помехи в спектре зачастую сопоставимы, в частности, из-за нелинейности усилительной аппаратуры, поэтому в исследованиях была произведена идентификация и верификация значений основной частоты путем раскадровки видеозаписи махов крыльями самцов и самок An. atroparvus, отснятой с помощью скоростной камеры «C100 Centurio» с частотой 1700–2000 кадров в секунду. В результате удалось достоверно сопоставить периодичность биения крыльев значениям их основной звуковой частоты. Основная частота и гармоники определяют индивидуальный тембр жужжания каждого насекомого. При этом установлено, что количество махов крыльями в единицу времени у самцов An. atroparvus значимо больше, чем у самок. Данная закономерность оказалась общей для разных полов всех четырех видов: самки жужжат достоверно «ниже», чем самцы этого же вида (p< 0,05). Очевидно, что различия частоты взмахов крыльями обусловлены, главным образом, разницей размеров и веса тела, геометрией и площадью крыльев.
Межвидовой сравнительный анализ акустики трех представителей Anopheles с учетом полов показал неоднозначные различия в значениях их частот.
У самок An. atroparvus в звуковом спектре выявлены три пика (рис. 3, Female): первый, самой высокой амплитуды – основная частота (среднее значение для выборки f = 293 Гц, число особей n = 72); второй и третий пики – гармоники, или обертоны, кратные значению главного пика (584 и 880 Гц, соответственно).
У самцов An. atroparvus два хорошо выраженных пика (третий слабо отличается от фоновых колебаний) (рис. 3, Male): первый (основная частота), среднее значение для выборки f = 444 Гц (n = 29); второй пик – гармоника – средняя частота 887 Гц. Необходимо подчеркнуть, что анализируемые частоты особей одного пола An. atroparvus статистически совпадают на уровне значимости α = 0,05. Таким образом, средняя основная частота самца выше, чем у самки, на 151 Гц, соответственно различаются и гармоники.
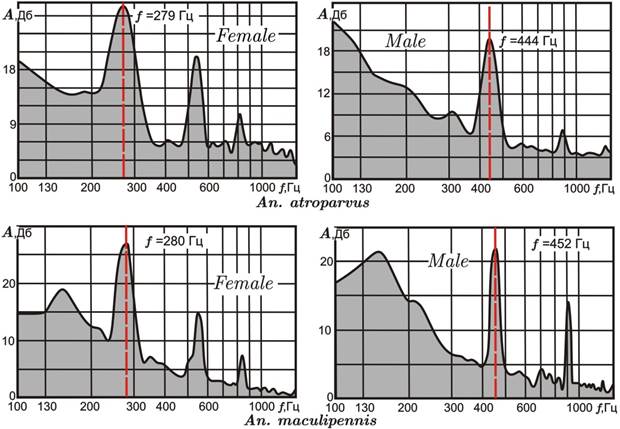
Рисунок 3. Типовые спектры звуковых частот самок (Female) и самцов (Male) An. atroparvusи An. maculipennis
Красными пунктирными линиями на диаграммах отмечены пики основных частот
У самок An. maculipennis в звуковом спектре выявлены также три пика (рис. 3, Female): первый (основная частота) – среднее значение для выборки f = 272 Гц; второй и третий (гармоники) – 548 и 820 Гц, соответственно (n = 11). У самцов An. maculipennis так же, как и у An. atroparvus, в звуковом спектре хорошо выражены два пика (рис. 3, Male): первый (основная частота) – среднее значение для выборки f = 448 Гц (n = 15); второй пик (гармоника) – средняя частота 890 Гц. Средняя основная частота самца выше, чем у самки, на 176 Гц, соответственно различаются и гармоники.
Таким образом, An. atroparvus и An. maculipennis имеют близкие акустические характеристики (у самцов они практически совпадают), чем в частности можно объяснять феномен межвидовой гибридизации этих двух видов. Единичные межвидовые гибриды на стадии личинок, отловленные ранее в природных биотопах Молдавии (г. Бельцы) и Калмыкии (г. Элиста), были выявлены посредством цитогенетического анализа [3, 7].
Обнаружение природных гибридов An. atroparvus × An. maculipennis является уникальным к настоящему времени событием среди всех видов комплекса «Anophelesmaculipennis». Данный феномен также подтверждается результатами скрещивания видов в лабораторных условиях, как путем свободной копуляции в садках, так и методом принудительной копуляции [6].
Лабораторная гибридизация An. atroparvus × An. maculipennis показала полную стерильность самцов F1 и некоторую фертильность самок F1. Последние, будучи способными к скрещиванию с самцами An. atroparvus, совершенно не развивали яиц. Очевидно, репродуктивная изоляция между An. atroparvus и An. maculipennis, четко проявляющаяся на уровне стерильности гибридов, не подкрепляется поведенческой изоляцией. Можно сделать вывод, что абсолютных этологических барьеров при скрещивании между этими видами не существует, что хорошо соотносится с полученными характеристиками их звуковых частот.
Акустические параметры третьего изученного вида – An. messeae – значимо отличаются от таковых первой описанной пары. Во-первых, учитываемые частоты An. messeae достоверно ниже, чем у An. atroparvus и An. maculipennis. Во-вторых, для самцов An. messeae установлены два типовых варианта пиков в спектре частот. В первом варианте, в отличие от An. atroparvus и An. maculipennis, наблюдаются три выраженных одинарных пика, так же как и у самок (рис. 4а и 4б). Во втором варианте для самцов с определенными кариотипами характерны парные («пакетные») пики обертонов (рис. 4в). Следует подчеркнуть, что гибриды от скрещивания An. messeae с An. atroparvus или с An. maculipennis в природе не обнаружены.
В-третьих, диапазон главных частот, прежде всего у самцов An. messeae, оказался гораздо разнообразнее, чем у других исследованных видов, что, очевидно, связанно с внутривидовым инверсионным полиморфизмом этого представителя. Перестройки затрагивают как половую хромосому (XL), так и плечи двух аутосом (2R, 3R, 3L) [9].
В юго-западных регионах Палеарктики преимущественно встречаются эволюционно исходные варианты – XL0 (XL1), 2R0, 3R0, 3L0; в северо-восточных – XL1, XL2, 2R1, 3R1, 3L1. В центре ареала An. messeae популяции характеризуются смешенным составом инверсий, хотя их определенные сочетания в кариотипах регистрируются значимо чаще, чем это ожидалось исходя из частот отдельных перестроек [4, 6]. В соответствие с этим было сделано предположение, что в полиморфных гемипопуляциях имаго An. messeae имеет место ассортативное скрещивание (предпочтительное спаривание генетически сходных особей).
Статистический анализ по критерию Стьюдента не выявил значимых различий по всем частотным пикам у самок с различными кариотипами (основная частота от 211 до 225 Гц).
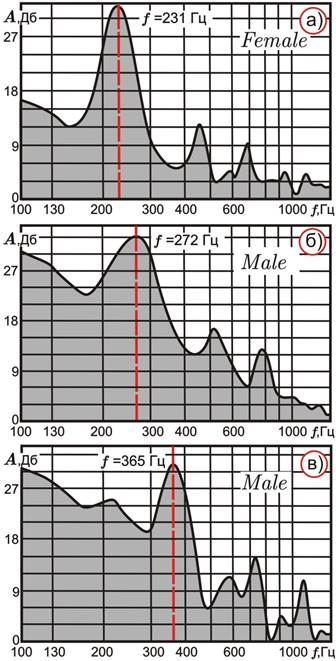
Рисунок 4. Типовые акустические спектры самки и самцов An. messeae
В то же время у самцов достоверные различия наблюдаются практически между всеми выделенными вариантами, особенно между имаго с кариотипами, распространенными или доминирующими в разных частях ареала (для «юго-западных» форм основная частота 261‑301 Гц, для «северо-восточных» – 326-363 Гц). Следует подчеркнуть, что эффекты ассоциативного взаимодействия инверсий в кариотипах наиболее выражено проявляются именно у самцов [6].
Известно, что самцы Anopheles в вечернее время собираются в рои, чтобы усилить звучание для привлечения самок [2]. Остается открытым вопрос, каким образом происходит окончательная идентификация самкой полового партнера, так как кариотипическая структура роев до сих пор не изучалась.
Возможны два варианта: либо рой преимущественно состоит из самцов с определенными кариотипами, и самка удаленно его идентифицирует; либо рой представляет собой полиморфную гемипопуляцию самцов, и тогда выбор самкой полового партнера осуществляется уже внутри роя.
Акустические характеристики имаго An. messeae указывают на то, что комары с более высокой частотой жужжания обитают в северных частях ареала, и, наоборот, «южные» формы характеризуются относительно низкими показателями. Можно предположить, что имаго, распространенные на севере ареала с коротким летом, имеют большие размеры, что позволяет откладывать самкам больше яиц и быстро наращивать численность популяций. «Южные» комары имеют меньшие размеры и плодовитость. В таком случае более крупные «северные» особи в полете должны более интенсивно махать крыльями (при равенстве других параметров), и, как следствие, иметь более высокие значения звуковых частот.
Особняком в комплексе «Anophelesmaculipennis» позиционируется вид An. beklemishevi: он имеет наиболее отдаленное родство с остальными видами и, очевидно, другой путь проникновения в Палеарктику из Северной Америки [6]. Данный представитель распространен по северной таежной части Евразии и является викарирующим с An. maculipennis в Европе и симпатриантом с An. mеsseae преимущественно в северной части ареала последнего [5]. Для An. beklemishevi установлены следующие акустические характеристики: средняя основная частота у самок – 253 Гц (и кратные ей 2 гармоники), у самцов – 269 Гц (и кратные ей 2 гармоники, как у An. messeae). Таким образом, вид отличается по своим частотам от всех остальных Anopheles. Обращает на себя внимание самое минимальное различие частот между полами (отношение средних частот самец/самка всего 1,06, тогда как у других: An. maculipennis – 1,51; An. mеsseae – 1,55; An. atroparvus – 1,52). Интересно также отметить, что наибольшие отличия по звуковым частотам An. beklemishevi проявляет по отношению к «северным» кариотипическим формам An. mеsseae, с которыми в большей степени совместно обитают. И наоборот, имаго An. messeae с наборами инверсий, распространенных на юго-западе ареала, наибольшим образом отличаются по акустике от других европейских изученных видов – An. atroparvus и An. maculipennis, что логичнее всего объяснить симпатрией An. messeae с этими видами именно на юго-западе Палеарктики.
Таким образом, полученные результаты акустических характеристик близкородственных видов малярийных комаров позволяют рассматривать их звуковую коммуникацию как основной механизм для видовой и внутривидовой идентификации полового партнера при спаривании.
Работа выполнена при поддержке Минобрнауки России в рамках базовой части государственного задания № 2014/387/1029.
Рецензенты:
Степанов В.А., д.б.н., профессор, заместитель директора института по научно-исследовательской работе, руководитель лаборатории эволюционной генетики ФГБНУ «Научно-исследовательский институт медицинской генетики», г. Томск;
Сибатаев А.К., д.б.н., старший научный сотрудник лаборатории эволюционной цитогенетики НИИ биологии и биофизики при ТГУ (НИИББ), г. Томск.
Библиографическая ссылка
Перевозкин В.П., Бондарчук С.С., Иваницкий А.Е., Петрова Н.В. АКУСТИЧЕСКИЕ ХАРАКТЕРИСТИКИ МАЛЯРИЙНЫХ КОМАРОВ КОМПЛЕКСА ANOPHELES MACULIPENNIS // Современные проблемы науки и образования. 2015. № 3. ;URL: https://science-education.ru/ru/article/view?id=17359 (дата обращения: 28.07.2026).