



В настоящее время интенсификация селекционного процесса в свиноводстве требует научно-обоснованных подходов в селекции. Необходимым условием повышения эффективности племенного отбора является получение точной информации о продуктивности животных еще в раннем возрасте, а также возможности использования полного его генетического потенциала [2, 5]. Общепринятые методы селекции свиней не всегда позволяют полностью использовать генетический потенциал существующих пород [7]. Поэтому в последние годы усилия отечественных и зарубежных специалистов направлены на поиск и внедрение инновационных методов, имеющих значительный потенциал наряду с традиционной селекцией. В качестве таких методов могут выступать технологии, основанные на исследованиях в области молекулярной генетики. Внедрение этих технологий в селекционно-племенную работу требует проведения исследований, прежде чем они могут быть эффективно использованы в свиноводстве [3].
Использование маркерной селекции приобретает большое значение в повышении воспроизводительных качеств свиней, так как их улучшение традиционными методами затруднено в силу низких значений коэффициентов наследуемости [4]. Перспективным приемом повышения воспроизводительных качеств свиней является использование ДНК-маркеров плодовитости [6].
На сегодняшний день большой интерес представляет ген лейкемия ингибирующего фактора (LIF) как генетический маркер воспроизводительных качеств свиней.
LIF был впервые идентифицирован в 1986 г. Metkalf et al. в исследованиях на мышах. На мышиных моделях впервые было показано участие цитокинов семейства IL-6 в имплантации, в частности в экспериментах с инактивацией гена LIF. Самки мышей с инактивированным геном LIF оказались бесплодными из-за нарушения имплантации бластоцисты [1].
Цитокины и факторы роста обладают способностью связываться со специфическими рецепторами клеточных поверхностей и выступают в качестве потенциальных внутриклеточных сигналов, регулирующих функции клеток эндометрия. В настоящее время у большинства видов эндометрий признан важнейшим местом образования цитокинов и рецепторов к ним [10].
Влияние LIF и его рецепторов было продемонстрировано на большом числе видов млекопитающих [8]. По данным A. Spotter et al. (2003), K. Ropka-Molik et al. (2012), Napierała D. et al (2014), ген LIF/DraIII может выступать как достаточно информативный генетический маркер воспроизводительных качеств свиней [8, 9, 10]. В ранее проведенных нами исследованиях на свиньях породы ландрас в ЗАО «Племзавод Юбилейный» Тюменской области было установлено влияние гена LIF/DraIII на воспроизводительные качеств свиней. Свиноматки генотипа АА/LIF по сравнению с аналогами генотипа ВВ/LIF достоверно отличались лучшими показателями по количеству поросят при рождении и многоплодию на 1,6 и 1,7 поросенка (р≤0,05), соответственно. Свиноматки генотипа АВ/LIF имели промежуточные значения. Полученные нами результаты свидетельствуют о том, что концентрация «желательного» аллеля А/LIF в генотипе животных способствует повышению многоплодия [1].
Наличие связи аллельных вариантов гена LIF/DraIII с плодовитостью свиней представляет интерес в изучение полиморфизма гена (SNP 6988 C→T) у свиней различных пород.
В связи с этим, целью данных исследований стало определение генетической структуры свиней различных пород по гену LIF.
Материалы и методика исследований
Исследования проводили на базе лаборатории молекулярной диагностики и биотехнологии с.-х. животных Донского государственного аграрного университета. Для определения генетической структуры свиней (n=507) по данным полиморфизма гена LIF в анализ были включены свиньи породы ландрас (n=326), свиньи крупной белой породы n=(135) и свиньи породы дюрок (n=46).
Для проведения молекулярно-генетических исследований у свиней были отобраны образцы ткани с ушной раковины (ушные выщипы) площадью 1 см².
Анализ проводили с помощью метода ПЦР-ПДРФ. Для проведения ПЦР использовали олигонуклеотидные праймеры (ЗАО «Евроген»): 5'-ATGTGGATGTGGCCTACGG-3' (GenBank AJ296176, nucleotide 6842-6861); 3'-GGGAACAAGGTGGTGATGG-5' (GenBank AJ296176, nucleotide 7231-7249).
Условия ПЦР: первоначальная денатурация - 4 мин при 940С; денатурация 940С - 30с, отжиг 580С - 60с, элонгация 720С - 30 с (30 циклов), завершающая элонгация при 720С 4 мин.
ПЦР-ПДРФ анализ фрагмента гена LIF длиной 407 н.п. проводили с использованием рестриктазы DraIII (ООО «СибЭнзим-М»). Рестрикционные фрагменты разделяли в 2%-ном агарозном геле с добавлением бромистого этидия. При визуализации электрофореграмм в УФ свете один фрагмент длиной 407 п.н. определяли как генотип АА, два фрагмента длиной 266 и 144 п.н. как генотип ВВ, три фрагмента длиной 407, 266 и 144 п.н. соответствовали генотипу АВ [1].
По результатам молекулярно-генетического исследования определяли наличие и частоту аллелей и генотипов по гену LIF.
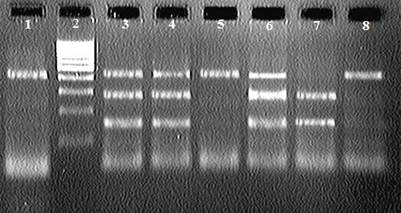
(Обозначения: 1 - ПЦР продукт (407 п.н.) ; 2 - ДНК-маркер 100 bp (СибЭнзим); 5,8 - генотип АА (407 н.п.); 7 - генотип ВВ (266- и 144 н.п.); 3,4,6 - генотип АВ (407-, 266- и 144 н.п.)).
Результаты исследований и их обсуждение
В результате проведения ДНК-генотипирования свиней по гену LIF была изучена генетическая структура свиней породы ландрас, крупная белая и дюрок. Данные результатов анализа представлены в таблице.
Частота аллелей и генотипов гена LIF свиней породы ландрас, крупной белой породы и породы дюрок
|
Порода |
Выборка,n |
Частота аллелей |
Частота генотипов, % |
||||||
|
А |
В |
AA |
AВ |
ВВ |
|||||
|
n |
% |
n |
% |
n |
% |
||||
|
Ландрас |
326 |
0,48 |
0,52 |
68 |
20,9 |
178 |
54,6 |
80 |
24,5 |
|
Крупная белая |
135 |
0,65 |
0,35 |
49 |
36,3 |
78 |
57,8 |
8 |
5,9 |
|
Дюрок |
46 |
0,24 |
0,76 |
4 |
8,8 |
14 |
30,5 |
28 |
60,7 |
Результаты анализа свиней породы ландрас по гену LIF/DraIII показали наличие двух аллелей А и B. В исследуемой популяции у свиней частоты аллелей А и В были распределены практически одинаково (0,48 и 0,52, соответственно). Низкой частотой обладал гомозиготный генотип АА (20,9%), а частота другого гомозиготного генотипа ВВ имела промежуточное значение (24,5%), частота гетерозиготного генотипа АВ составила 54,6%.
Генетическая структура свиней крупной белой породы по гену LIF/DraIII так же определяется наличием всех трех генотипов АА, АB и BB (таблица). В данной популяции наименьшей частотой обладал гомозиготный генотип BB (5,9%), частота генотипа АA имела промежуточное значение (36,3%). В исследуемой популяции, при оценке распределения частот, приоритетом располагал аллель А (0,65). Наибольшую частоту имел генотип АВ (57,8%).
Анализ частот аллелей и генотипов свиней породы дюрок (таблица) показал наличие трех генотипов гена LIF. В исследуемой популяции наибольшую частоту имел аллель B (76,1%) и генотип ВВ (60,9%). Наименьшая частота определена для генотипа АА (8,8). Анализ распределения частот аллелей свиней различных пород показывает, повышенную частоту аллеля В у свиней породы дюрок по сравнению со свиньями породы ландрас и крупной белой породы. Вероятно, такое распределение связано с направлением продуктивности данных свиней.
Следует отметить, что данные по изучению распределения частот аллелей и генотипов гена LIF/DraIII, а также его влияния на продуктивные качества свиней, в отечественной литературе отсутствуют (согласно литературным источникам в открытом доступе), но имеются данные зарубежных ученых, проводивших аналогичные исследования.
По данным результатов исследований A. Spötter et al. (2003) на свиньях породы немецкий ландрас частоты аллелей А и В составили 0,56 и 0,44, соответственно. В популяции преобладал гомозиготный генотип АВ (0,54), частоты гомозиготных генотипов АА и ВВ составили 0,34 и 0,12 [10] соответственно, что согласуется с полученным нами результатам.
Результаты, полученные A. Spötter (2003) при изучении генетической структуры свиней немецкой крупной белой породы (n=18) по гену LIF, показали низкую частоту аллеля А (0,25), что сказалось на распределении генотипов, где наиболее распространенным был генотип BB (0,51) и практически не встречался генотип AA (0,06) [10]. Данные результаты отличаются от полученного нами распределения у крупной белой породы.
Аналогичные результаты были получены в исследованиях A. Spötter et al. (2003) на свиньях породы дюрок, результаты которых показали наименьшую частоту аллеля А (0,33) [10], что согласуется с полученными нами результатами.
Выводы
Таким образом, на основании собственных исследований и литературных данных можно заключить, что для изучаемых пород характерны все три генотипа (АА, АВ и ВВ) гена LIF, но наибольшая частота, как правило, принадлежит гетерозиготному генотипу.
Сопоставляя частоты аллелей гена LIF у исследованного нами поголовья свиней c литературными данными, следует отметить породный и, возможно, географический аспект распределения частот аллелей и генотипов данного гена. Полученные результаты расширяют современные представления о молекулярных маркерах, характеризующих воспроизводительные качества свиней.
Рецензенты:
Приступа В.Н., д.с.-х.н., профессор кафедры частной зоотехнии и кормления сельскохозяйственных животных ФГБОУ ВПО ДГАУ, п. Персиановский;
Третьякова О.Л., д.с.-х.н., зав. лабораторией теоретических основ селекции с.-х. животных, профессор кафедры частной зоотехнии и кормления сельскохозяйственных животных ФГБОУ ВПО ДГАУ, п. Персиановский.
Библиографическая ссылка
Леонова М.А., Гетманцева Л.В., Колосов А.Ю. РАСПРЕДЕЛЕНИЕ ЧАСТОТ АЛЛЕЛЕЙ И ГЕНОТИПОВ ГЕНА ЛЕЙКЕМИЯ ИНГИБИРУЮЩЕГО ФАКТОРА У СВИНЕЙ РАЗЛИЧНЫХ ПОРОД // Современные проблемы науки и образования. 2015. № 2-1. ;URL: https://science-education.ru/ru/article/view?id=17343 (дата обращения: 26.06.2026).