



Изучение генетического разнообразия и внутривидовой дифференциации лесообразующих видов хвойных, в том числе и видов рода Larix, имеющих важное биосферное и ресурсное значение, является одной из важнейших задач популяционной биологии. Только на основании точных сведений о генетической структуре популяций хвойных, уровне их генетической изменчивости и характере ее распределения в пределах ареалов видов может быть оценен генетический потенциал видов и разработан для каждого из них комплекс мероприятий, направленных на максимальное сохранение генетического разнообразия в процессе их использования и воспроизводства [1]. Генетическое разнообразие лиственницы сибирской на Урале изучалось В.Л. Семериковым на основании полиморфизма цитоплазматических маркеров и частично ядерных генов [7]. Однако полиморфизм межмикросателлитных молекулярных маркеров в геноме этого вида до настоящего времени не был исследован, в том числе в популяциях L. sibirica Пермского края.
Цель данной работы – выявление полиморфизма ISSR-PCR маркеров, параметров генетического разнообразия и генетической структуры семи популяций лиственницы сибирской на Урале.
Материал и методы
В 2009-2014 гг. было изучено семь популяций Larix sibirica Ledeb (семейство Pináceae). Шесть из изученных популяций находятся в Пермском крае: две популяции являются природными и расположены в равнинных областях (Ls1 около пос. Полазна в центральной части края, Ls2 около г. Оса на юго-западе края); две популяции являются искусственными насаждениями (Ls4 на территории ООПТ «Парковый» Очерского района в западной части Пермского края, заложены известным лесоведом Ф.А. Теплоуховым; Ls5 в Кишертском районе на территории учебно-научной базы «Предуралье» в восточной части края); две популяции расположены на северо-востоке края в заповеднике «Вишерский» в горно-лесном поясе западного склона Уральских гор (Ls6 на западном склоне хребта Тулымский камень, Ls7 на правом берегу р. Таборная). В Свердловской области около пос. Билимбай в зоне низкогорной тайги находится еще одна природная популяция L. sibirica (Ls3). Она интересна тем, что расположена с восточной стороны Уральских гор.
Для молекулярно-генетического анализа в каждой популяции была собрана хвоя с 28-32 деревьев примерно одного возраста, расстояние между деревьями составляло не менее 80 м. Наименьшее расстояние между популяциями – 50 км, среднее расстояние между популяциями – 200 км. Полиморфизм ДНК проанализирован у 205 отдельных деревьев. ДНК выделяли из 20 мг высушенной хвои с использованием СТАВ-метода с добавлением в качестве сорбента PVPP (polyvinylpolypyrrolidone) [5]. Концентрацию и спектральные характеристики ДНК определяли на спектрофотометре NanoDrop 2000 (Thermo scientific, USA). Тотальная ДНК была разбавлена до рабочей концентрации 10 нг/мкл. Молекулярно-генетический анализ проведен с использованием ISSR (Inter-Simple Sequence Repeats) - метода анализа полиморфизма ДНК [10]. Анализ эффективности произведен у 20 ISSR-праймеров по методике, предложенной Р.Н. Календарем и С.В. Боронниковой [2] по шкале от 1 (низкая) до 5 (самая высокая). Реакционная смесь для ISSR-ПЦР объемом 25 мкл содержала: 2 единицы Tag-полимеразы, 2,5 мкл стандартного 10х ПЦР-буфера, 25 пМ праймера, 2,5 мМ Mg2+, 0,25 мM dNTP, 5 мкл геномной ДНК. Амплификацию проводили в термоциклере GeneAmp PCRSystem 9700 (Applied Biosystems, USA) по типичной для ISSR-метода программе: предварительная денатурация 94 °C, 2 мин.; первые пять циклов 94 °С, 20 сек.; t° отжига, 10 сек.; 72 °С, 10 сек.; в последующих тридцати пяти циклах 94 °С, 5 сек.; tо отж., 5 сек.; 72 °С, 5 сек. Последний цикл элонгации длился 2 мин при 72 ºС. Температура отжига в зависимости от G/С-состава праймеров варьировала от 56 до 64 °С. В качестве отрицательного (К-) контроля в реакционную смесь для проверки чистоты реактивов добавляли вместо ДНК 5 мкл деионизированной воды. Продукты амплификации разделяли электрофорезом в 1,7%-ном агарозном геле. Для определения длины фрагментов ДНК использовали маркер молекулярной массы (100bp+1.5+3Кb DNA Ladder, ООО «СибЭнзим-М», Москва). Компьютерный анализ полиморфизма ДНК проведен с помощью программы POPGENE 1.32 и специализированного макроса GenAlEx6 для MS-Excel. Для описания генетической структуры популяций L. sibirica были использованы следующие параметры [8]: ожидаемая доля гетерозиготных генотипов (HT) во всей популяции как мера общего генного разнообразия; ожидаемая доля гетерозиготных генотипов (HS) в субпопуляции как мера ее внутрипопуляционного разнообразия; доля межпопуляционного генетического разнообразия в общем разнообразии или показатель подразделенности популяций (GST). Генетическое расстояние (D) между популяциями определено по формуле M. Нея и В. Ли [9].
Результаты и их обсуждение
Каждый из 20 ISSR-праймеров был индивидуально проанализирован в ПЦР с геномной ДНК L. sibirica. В результате для данного вида отобраны 5 эффективных ISSR-праймеров, они выявляли наибольшее число четко амплифицирующихся и стабильно воспроизводящихся фрагментов ДНК: M3 [(AC)8CT], M9 [(ACC)6G], X10 [(AGC)6C], ISSR-8 [(GAG)6C], CR-215 [(CA)6GT]. Из них 2 праймера являются динуклеотидными, а 3 праймера - тринуклеотидными, так называемые якорные нуклеотиды отмечены как одно-, так и двунуклеотидные.
В семи популяциях L. sibirica было детектировано 119 ISSR-PCR маркеров, из которых 116 были полиморфными (![]() =0,974). В среднем один ISSR-праймер инициировал у L. sibirica синтез 17,1 фрагментов ДНК (рис. 1).
=0,974). В среднем один ISSR-праймер инициировал у L. sibirica синтез 17,1 фрагментов ДНК (рис. 1).
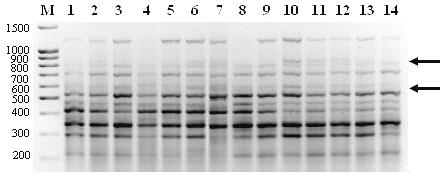
Рис. 1. ISSR-спектр популяции Ls1 с праймером Х10. М – маркер молекулярного веса,
1-4 – номера проб ДНК, стрелками обозначены некоторые полиморфные фрагменты.
Доля полиморфных локусов выше в популяции Ls5 (![]() =0,830), ниже в Ls2 (
=0,830), ниже в Ls2 (![]() =0,479). Из 119 ISSR-маркеров 14 (11,8%) являются уникальными, они представлены только в одной популяции, а 105 ISSR-маркеров (88,2%) являются общими для всех семи исследованных популяций. При сравнении доли полиморфных локусов по критерию Фишера популяция Ls5 имеет значимые отличия от всех остальных популяций (F>Fst=1,96). Также значимая разница по этому показателю обнаружена между популяцией Ls2, популяциями Ls1 и Ls4. Самой распространенной мерой генетической изменчивости в популяции является гетерозиготность. Ожидаемая гетерозиготность (
=0,479). Из 119 ISSR-маркеров 14 (11,8%) являются уникальными, они представлены только в одной популяции, а 105 ISSR-маркеров (88,2%) являются общими для всех семи исследованных популяций. При сравнении доли полиморфных локусов по критерию Фишера популяция Ls5 имеет значимые отличия от всех остальных популяций (F>Fst=1,96). Также значимая разница по этому показателю обнаружена между популяцией Ls2, популяциями Ls1 и Ls4. Самой распространенной мерой генетической изменчивости в популяции является гетерозиготность. Ожидаемая гетерозиготность (![]() ) на общую выборку составила 0,125 (табл. 1). Этот показатель наибольший в популяции Ls5 (HE = 0,168), а наименьший – в популяции Ls2 (HE =0,074), данные значения достоверно отличаются (F=2,149>Fst=1,96) при использовании критерия Фишера.
) на общую выборку составила 0,125 (табл. 1). Этот показатель наибольший в популяции Ls5 (HE = 0,168), а наименьший – в популяции Ls2 (HE =0,074), данные значения достоверно отличаются (F=2,149>Fst=1,96) при использовании критерия Фишера.
Таблица 1
Генетическое разнообразие изученных популяций L. sibirica
|
Популяции/ показатели |
Ls1 |
Ls2 |
Ls3 |
Ls4 |
Ls5 |
Ls6 |
Ls7 |
На выборку |
|
P95 |
0,605 |
0,479 |
0,554 |
0,638 |
0,830 |
0,558 |
0,487 |
0,974 |
|
HE |
0,121 (0,117) |
0,074 (0,013) |
0,124 (0,017) |
0,157 (0,017) |
0,168 (0,015) |
0,135 (1,018) |
0,093 (0,015) |
0,125 (0,008) |
|
na
|
1,369 (0,484) |
1,268 (0,445) |
1,352 (0,479) |
1,470 (0,501) |
1,630 (0,484) |
1,386 (0,489) |
1,285 (0,453) |
2,000 (0,000) |
|
ne
|
1,207 (0.341) |
1,119 (0.257) |
1,208 (0,326) |
1,260 (0,340) |
1,256 (0.285) |
1,232 (0,349) |
1,159 (0,308) |
1,442 (0,321) |
|
R |
2 |
0 |
0 |
1 |
10 |
0 |
1 |
14 |
Примечание: P95 – доля полиморфных локусов, HE – ожидаемая гетерозиготность, na – абсолютное число аллелей на локус; ne - эффективное число аллелей на локус, в скобках даны стандартные отклонения; R – число редких аллелей; жирным шрифтом выделены максимальные, а подчеркиванием – минимальные значения.
Для характеристики генетической структуры популяций важны также редкие, то есть встречающиеся с частотой менее 5% маркеры. В изученных популяциях L. sibirica выявлено 14 редких ISSR-PCR маркеров. Наибольшее число редких аллелей (R =10) отмечено в популяции Ls5 (рис. 2).
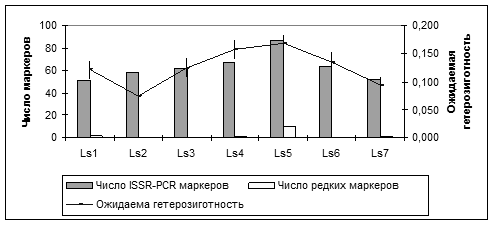
Рис. 2. Выявленные ISSR-PCR маркеры и ожидаемая гетерозиготность в изученных популяциях L. Sibirica.
Абсолютное число аллелей выше (табл. 1) в популяциях Ls5 (![]() =1,630) и Ls4 (
=1,630) и Ls4 (![]() =1,470). Эффективное число аллелей оказалось наибольшим в популяции Ls4 (
=1,470). Эффективное число аллелей оказалось наибольшим в популяции Ls4 (![]() =1,260) и близко по значению в популяции Ls5 (
=1,260) и близко по значению в популяции Ls5 (![]() =1,256). Этот показатель достоверно ниже в двух популяциях Ls2 (
=1,256). Этот показатель достоверно ниже в двух популяциях Ls2 (![]() =1,119) и Ls7 (
=1,119) и Ls7 (![]() =1,159). Полученные нами данные согласуются с приведенными в литературе показателями генетического полиморфизма по ISSR-PCR маркерам для хвойных видов растений. Так, для ели гибридной доля полиморфных локусов в популяциях составляет 0,970, абсолютное и эффективное число аллелей также близки к таковым для лиственницы сибирской (na=1,970 и ne=1,463) [6]. У сосны обыкновенной [3; 4] отмечена меньшая доля полиморфных локусов (0,940) по сравнению с изученными нами популяциями L. sibirica. Эффективное число аллелей в популяциях сосны обыкновенной (ne=1,405) близко к значениям, установленным для других хвойных видов растений.
=1,159). Полученные нами данные согласуются с приведенными в литературе показателями генетического полиморфизма по ISSR-PCR маркерам для хвойных видов растений. Так, для ели гибридной доля полиморфных локусов в популяциях составляет 0,970, абсолютное и эффективное число аллелей также близки к таковым для лиственницы сибирской (na=1,970 и ne=1,463) [6]. У сосны обыкновенной [3; 4] отмечена меньшая доля полиморфных локусов (0,940) по сравнению с изученными нами популяциями L. sibirica. Эффективное число аллелей в популяциях сосны обыкновенной (ne=1,405) близко к значениям, установленным для других хвойных видов растений.
При сравнении трех групп популяций L. sibirica (искусственные насаждения, природные равнинные популяции, природные горные и предгорная популяции) доля полиморфных локусов достоверно выше в группе популяций искусственного происхождения (P95=0,848) по сравнению с группами природных популяций (F=4,8>Fst=1,96). Однако другие показатели генетического разнообразия (HE =0,223; ne =1,368) выше в группе горных и предгорной популяций данного вида (табл. 2). Значимо наиболее низкие показатели генетического разнообразия установлены в природных популяциях, расположенных в равнинных частях Пермского края (P95=0,563; HE =0,169; ne =1,284).
Таблица 2
Генетическое разнообразие трех групп популяций L. sibirica
|
Популяции/ показатели |
Природные равнинные (Ls1, Ls2) |
Искусственные насаждения (Ls4, Ls5) |
Природные горные и предгорная (Ls3, Ls6, Ls7) |
|
P95 |
0,563 |
0,848 |
0,756 |
|
HE |
0,169 (0,018) |
0,214 (0,014) |
0,223 (0,017) |
|
ne |
1,284 (0,352) |
1,329 (0,289) |
1,368 (0,353) |
Примечание: P95 – доля полиморфных локусов, HE – ожидаемая гетерозиготность, na – абсолютное число аллелей на локус; ne - эффективное число аллелей на локус, в скобках даны стандартные отклонения; жирным шрифтом выделены максимальные, а подчеркиванием – минимальные значения.
Анализ генетической структуры изученных популяций L. sibirica показал, что ожидаемая доля гетерозиготных генотипов (HT) на общую выборку составила 0,273, а ожидаемая доля гетерозиготных генотипов в отдельной популяции по всем локусам (HS) равна 0,125. Коэффициент подразделенности популяций показывает, что на межпопуляционную компоненту приходится 54,35% всего генетического разнообразия (табл. 3). Наибольшая дифференциация между популяциями L. sibirica установлена с использованием праймера X10.
Таблица 3
Генетическая структура изученных популяций L. sibirica
|
ISSR-праймер |
HT |
HS |
GST |
|
M3 M9 Х10 ISSR-8 CR-215 |
0,285 (0,023) 0,308 (0,025) 0,249 (0,036) 0,296 (0,030) 0,265 (0,035) |
0,162 (0,012) 0,147 (0,006) 0,080 (0,005) 0,130 (0,006) 0,117 (0,006) |
0,432 0,521 0,681 0,559 0,557 |
|
На общую выборку |
0,273 (0,024) |
0,125 (0,008) |
0,543 |
Примечание: HT – общее генное разнообразие; HS – внутрипопуляционное разнообразие; GST – показатель подразделенности популяций; в скобках даны стандартные отклонения.
Наименьшее генетическое расстояние отмечено между популяциями Ls6 и Ls7 (D=0,105), а наибольшее – между популяциями Ls2 и Ls7 (D=0,414).
Заключение
Изученные семь популяций L. sibirica характеризуются высоким уровнем генетического разнообразия на основании полиморфизма ISSR-маркеров. Доля полиморфных локусов в общей выборке составила 97,4%, а ожидаемая гетерозиготность - 0,125. Из семи изученных популяций самые высокие показатели генетического разнообразия отмечены в искусственных насаждениях Ls5 (P95 = 0,830; HE = 0,168; ne = 1,256; R = 10) и Ls4 (P95 = 0,638; HE = 0,157; ne = 1,260; R =1), а самые низкие - в природной популяции Ls2 (P95 = 0,479; HE = 0,074; ne = 1,119; R = 0). Наибольшее число редких аллелей (R =10) отмечено также в популяции Ls5. Полученные результаты согласуются с литературными данными о полиморфизме ISSR-PCR маркеров для других видов хвойных растений. Генетически наиболее близки природные горные популяции Ls6 и Ls7 (D=0,105), расположенные на меньшем географическом расстоянии, наиболее генетически далеки популяции Ls2 и Ls7 (D=0,414). Исследованные популяции L. sibirica значительно дифференцированы, в связи с чем для сохранения генофонда необходимо выбирать популяции из разных частей ареала с учетом происхождения, а также географических и экологических условий произрастания популяций.
На основании результатов исследований даны следующие рекомендации.
1. Для сохранения и возобновления генетических ресурсов L. sibirica на Урале рекомендуется использовать искусственные насаждения лиственницы сибирской Ls5 (P95 =0,830; HE =0,168; ne =1,256; R =10) и Ls4 (P95 =0,638; HE =0,157; ne =1,260; R =1), заложенные еще Ф.А. Теплоуховым; одну популяцию, расположенную в центральной равнинной части Пермского края Ls1 (P95 =0,605; HE =0,121; ne =1,207; R=2); а также популяцию из горно-лесного пояса Урала Ls6 (P95 =0,558; HE =0,135; ne =1,232; R=0).
2. При отборе деревьев для лесовосстановления необходимо учитывать их генотип, а также наличие редких ISSR-PCR маркеров, которые наряду с другими молекулярными маркерами могут быть использованы и для молекулярно-генетической идентификации популяций.
3. Для сохранения генетического разнообразия лиственницы сибирской на популяционном уровне необходимо учитывать генетическую структуру популяций и уровень внутри- и межпопуляционной дифференциации.
Работа выполнена при финансовой поддержке задания 2014/153 на оказание государственных работ в сфере научной деятельности в рамках базовой части государственного задания Минобрнауки России (№ гос. регистрации 01201461915).
Рецензенты:
Янбаев Ю.А., д.б.н., профессор, проректор по учебной работе ФГБОУ ВПО «Башкирский государственный университет», г. Уфа.
Плотникова Е.Г., д.б.н., ведущий научный сотрудник ФГБОУН «Институт экологии и генетики микроорганизмов Уральского отделения Российской академии наук» (ИЭГМ УрО РАН), г. Пермь.
Библиографическая ссылка
Нечаева Ю.С., Боронникова С.В., Пришнивская Я.В., Чумак Е.И., Юсупов Р.Р., Нечаева Ю.С., Боронникова С.В., Пришнивская Я.В. АНАЛИЗ ПОЛИМОРФИЗМА ISSR-PCR МАРКЕРОВ И ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ НЕКОТОРЫХ ПОПУЛЯЦИЙ ЛИСТВЕННИЦЫ СИБИРСКОЙ НА УРАЛЕ // Современные проблемы науки и образования. 2014. № 6. ;URL: https://science-education.ru/ru/article/view?id=16047 (дата обращения: 29.05.2026).