



Самый северный в мире остров леса Ары-Мас на полуострове Таймыр лежит в долине правого притока Хатанги – р. Новой на широте 72°30´, далеко за пределами полярной границы лесов. Ары-Мас изолирован от прихатангских редколесий полосой кустарниковых тундр. Несмотря на то, что области контакта леса и тундры посвящены многочисленные исследования [1–7], отдельные аспекты их взаимоотношений до сей поры выяснены не полностью.
Известный специалист в области ботанической географии А.И. Толмачев писал о редколесьях из лиственницы к востоку от р. Пясины: «Хотя и есть местная деградация, крайняя жизненность характерна для них в условиях северной границы: обилие молодняка даже в области господства тундры, возможно, свидетельствует о распространении лиственничных насаждений на новые территории» [4].
Первый исследователь острова леса Ары-Мас Л.Н. Тюлина [7] отметила, что описанные А.И. Толмачевым характеристики свойственны и исследованным ею редколесьям Хатангского района. Кроме того, характерно равномерное распределение прихатангского редколесья по всем элементам рельефа почти у самой границы с тундрой, следовательно, лиственница растёт здесь в разнообразных условиях, а не только в наилучших. Почти во всех участках редколесья наряду с корявыми старыми экземплярами имеются вполне нормально развитые лиственницы более молодого поколения, растущие обычно гуще и местами уже образующие господствующий полог. Обильного сухостоя у границы нет. На основании данных исследований в Хатангском районе Л.Н. Тюлина высказала предположение о том, что лесная растительность здесь имеет тенденцию к продвижению в зону тундры.
Изменение соотношения между редколесьем и рединами в лесотундре наблюдается и в настоящее время. Целями наших исследований стало выявление определяющих динамику этого соотношения природных факторов и оценка роли отдельных климатических факторов в формировании годичного прироста стволовой древесины лиственницы.
Материалы и методы
В июле 2010 г. в лиственничных рединах и редколесьях лесного острова Ары-Мас были заложены две пробные площади по 0,25 га, на которых установлены автоматические температурные датчики, а также отобраны по 10 буровых кернов древесины из стволов лиственницы на каждой площади. Объектом изучения служила лиственница Гмелина (Larix gmelinii (Rupr.)).
Редколесья и редины выделены в ходе проведённого сотрудниками Института леса им В.Н. Сукачева СО РАН в 1985-1986 гг. лесоустройства по следующим параметрам: редколесья – древостои с полнотой 0,1-0,3; редины – древостои с полнотой 0,05–0,1. Высота деревьев в редколесьях составляет 4-6 м (до 11 м), диаметр стволов на высоте 1,3 м – 6-12 см (до 14–20 см в наилучших условиях), запас стволовой древесины на 1 га равен 5-10 м3.
Полученные буровые керны древесины были обработаны в Сибирском институте физиологии и биохимии растений СО РАН с применением автоматизированной системы LINTAB в программном пакете TSAP v. 3.5 [10]. После измерения ширины годичных колец были построены индивидуальные древесно-кольцевые хронологии, которые перекрестно датировались методом cross-dating. Затем датировка была проверена тестированием в программе COFECHA из пакета DPL [8]. Ряды индексов усреднялись для совокупности деревьев с одного участка, в итоге чего была получена стандартная обобщенная хронология [9].
Результаты и обсуждение
Лиственница Гмелина является на востоке Северо-Сибирской низменности единственной лесообразующей породой. Для лесов этого региона характерно участие представителей арктической и горной флоры, заметно ослаблена эдификаторная роль древесного яруса [2]. Редколесья с сомкнутостью крон 0,2–0,3 приурочены к нижней части склонов и выположенным участкам первой надпойменной террасы р. Новая. Они характеризуются развитым травяно-кустарничковым и кустарниковым ярусом, в котором доминируют багульниковые лесные ассоциации с развитым моховым покровом. Лиственничные редины отличаются от редколесий, прежде всего, геоморфологическим уровнем (вторая надпойменная терраса). Это верхний предел древесной растительности в урочище Ары-Мас. В рединах доминирует молодое поколение, а доля средневозрастных деревьев не превышает 30%. В целом для лесных растительных сообществ района исследований характерны изреженность древесного полога и низкая продуктивность древостоев, доминирование типов леса с мохово-лишайниковым, кустарниковым и кустарничковыми покровами, своеобразное сочетание элементов бореальной и тундровой флоры [1].
Территория относится к области распространения сплошных вечномёрзлых грунтов, мощность залегания которых превышает 300 м. Интенсивное развитие эрозионных процессов (термокарст, солюфликация, активность склоновых процессов и др.) обусловлено сезонным протаиванием почв. Глубина сезонного протаивания почвогрунта на северном пределе произрастания лиственницы в урочище Ары-Мас не превышает 50–70 см на минерализованных участках и 10–30 см под мощным моховым покровом [6].
Природные особенности района исследований обусловили крайне высокую зависимость древесной растительности от изменчивости ведущих климатических факторов – температуры воздуха и количества атмосферных осадков [6, 9].
Как было ранее установлено, на протяжении XX в. в лиственничниках Ары-Маса чётко выделяются две волны возобновления [9]. Первое массовое возобновление лиственницы произошло в середине 1950-х, а второе– в первой половине 1980-х гг. (рис.1). В указанные периоды при синхронном увеличении температуры и уровня атмосферного увлажнения создались условия для появления подроста, которые были успешно реализованы. Напротив, в периоды синхронного понижения температуры и уменьшения атмосферных осадков во второй половине 1950-х и 1970-х гг. темпы лесовозобновления резко снижались.
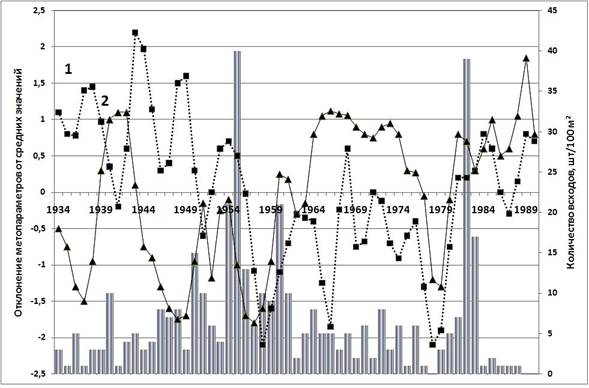
Рис. 1. Диаграмма динамики возобновления лиственницы в урочище Ары-Мас на фоне динамики температуры воздуха (1) и уровня атмосферного увлажнения (2)
Эти периоды были чётко зафиксированы и в динамике ширины годичных колец лиственницы как в рединах, так и в редколесьях. В то же время при анализе динамики радиального прироста деревьев в этих двух различных местообитаниях выяснились существенные различия. На рис. 2 представлены данные динамики ширины годичных колец лиственниц из редин и редколесий. Видно, что до середины 1950-х гг. наибольшим приростом характеризовались древостои редколесий, а со второй половины 1950-х гг. и до настоящего времени максимальный прирост наблюдается в рединах. На фоне чёткой погодичной синхронности сравниваемых древостоев не менее важно отметить разнонаправленность трендовых составляющих. Таким образом, можно утверждать, что, начиная с середины ХХ в., прирост деревьев в рединах и редколесьях различен. Мы вправе допустить, что в середине прошлого века произошла достаточно радикальная смена регулирующего климатического фактора, что и вызвало, прямым или косвенным образом, изменение динамики.
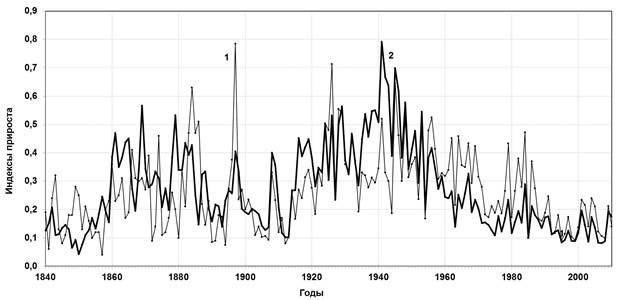
Рис. 2. Многолетняя динамика прироста ширины годичных колец древесины лиственницы Гмелина в рединах (1) и редколесьях (2) урочища Ары-Мас
Лесорастительные условия определяются комплексом взаимосвязанных факторов и процессов. Основным, определяющим во многом и действие других факторов, является баланс тепла и влаги. Существенное возрастание количества осадков в зимний период, а вместе с тем и мощности снегового покрова, меняет этот баланс. В зоне распространения вечной мерзлоты снежный покров важен не только как источник влаги в весеннее время, но и как теплоизолирующий экран, который предохраняет почву от чрезмерного охлаждения. В.В. Рахманов [6] отмечает различия характеристик снежного покрова на открытых местностях (поля, вырубки) и лесных участках. На территории наших исследований также можно условно выделить открытые и закрытые пространства, если за таковые соответственно принять редины и редколесья.
В апреле 2012 г. в период максимального снегонакопления на Ары-Масе мы провели снегомерные наблюдения в районе дендрохронологических исследований. На закрытых пространствах (в редколесьях) мощность снегового покрова составила 76,7 см, а плотность снега достигала 0,29 г/см3. В рединах средняя толщина снегового покрова была 27,5 см, при плотности 0,19 г/см3. Как видно, толщина снежного покрова в редколесьях почти втрое выше таковой в рединах при трехкратно большей плотности снега. Известно, что в лесу по сравнению с открытой местностью длительность снеготаяния напрямую зависит от количества выпавших за зиму осадков и плотности снежного покрова [2]. Учитывая, что продолжительность вегетационного периода на территории очень невелика и составляет 32-65 дней, каждый потерянный для вегетации день вносит существенный негативный вклад в продукционный процесс деревьев. Таким образом, большое количество осадков в зимний период не всегда можно считать благом. В случае позднего схода снежного покрова (в случае редколесий) вегетационный период деревьев начинается позднее, что приводит к снижению величин радиального прироста. Дополнительным негативным моментом может являться чрезмерная увлажненность деятельного слоя почвы при таянии мощного снежного покрова в плохо дренируемых местоположениях.
Ещё одной косвенной причиной, вызвавшей различия в радиальном приросте деревьев, может также являться различная мощность мохового покрова в редколесьях и рединах. Меньшая мощность мохового покрова в рединах обусловливает его более слабое термоизолирующее влияние и лучшую прогреваемость верхних слоёв почвы, а значит улучшает условия для развития корневой системы и обеспечения древесных растений элементами питания. В редколесьях же при более развитом моховом покрове уровень мерзлоты находится ближе к корнеобитаемому слою, что ухудшает условия питания. Глубина сезонного протаивания почвы в урочище Ары-Мас не превышает 50-70 см на минерализованных участках и 10–30 см под мощным моховым покровом [1].
Заключение
Таким образом, в результате проведённых исследований мы установили существенное изменение структуры атмосферного увлажнения района исследований в последние 50-60 лет. В результате роста количества осадков и внутригодового их перераспределения произошло заметное увеличение мощности снегового покрова, особенно в редколесьях лиственницы. В то же время заметного изменения термического режима в этот период не отмечено.
На протяжении XX в. в лиственничниках Ары-Маса дважды происходило массовое возобновление лиственницы: первое в середине 1950-х, а второе – в первой половине 1980-х гг. Эти периоды чётко зафиксированы и в динамике ширины годичных колец лиственницы в рединах и редколесьях (притом, что она существенно отличается в обоих местообитаниях). До середины 1950-х гг. наибольшим приростом характеризовались древостои редколесий, а со второй половины 1950-х годов xx в. и до настоящего времени максимальный прирост наблюдается в рединах. Наиболее вероятной причиной этого стало увеличение мощности снегового покрова в редколесьях и более позднее его разрушение по сравнению с рединами, из-за чего стартовые условия для ксилогенеза в раннелетний период здесь существенно ухудшились.
Дополнительным биотическим фактором, усугубляющим продукционный процесс в редколесьях, стал хорошо развитый моховой покров, препятствующий глубокому сезонному оттаиванию почвы. Произошла синергия негативных влияний позднего схода снега и неглубокого оттаивания корнеобитаемого слоя почвы. Это обстоятельство уменьшило продолжительность периода физиологической активности лиственницы и привело к зафиксированному снижению величины годичного радиального прироста древесины лиственницы в редколесьях по сравнению с рединами.
Рецензенты:
Воронин В.И., д.б.н., заместитель директора по научной работе федерального государственного бюджетного учреждения науки «Сибирский институт физиологии и биохимии растений» Сибирского отделения Российской академии наук (СИФИБР СО РАН), г. Иркутск;
Моложников В.Н., д.б.н., профессор, председатель Байкальского отдела Русского географического общества, г. Иркутск.
Библиографическая ссылка
Рудинский М.Г. ВОЗОБНОВЛЕНИЕ ЛИСТВЕННИЦЫ НА СЕВЕРНОМ ПРЕДЕЛЕ РАСПРОСТРАНЕНИЯ (МАССИВ АРЫ-МАС, П-ОВ ТАЙМЫР) // Современные проблемы науки и образования. 2014. № 6. ;URL: https://science-education.ru/ru/article/view?id=15687 (дата обращения: 03.07.2026).