



Введение
Оценка селекционного материала на основе белкового маркера дает возможность достаточно быстро и качественно проводить отбор и контролировать передачу желаемых признаков от родительских форм в гибридные популяции. Вместе с тем, для селекции необходимо выявление новых и стабильных белковых маркеров для определения качества зерна пшеницы. Наиболее широко изучаемыми белками пшеницы являются запасные белки глиадин и глютенин. Полная характеристика отдельных фракций и компонентов белков отражается во многих научных исследованиях. Поэтому изучение генетической закономерности накопления белков в зерне различных сортов пшеницы и использование при идентификации генотипов пшеницы в роли маркера компонент глиадиновых белков является наиболе актуальной проблемой современности [1].
Практически ряд молекулярных маркеров используется при исследовании пшеницы. Вместе c тем, среди изученных молекулярных маркеров более досконально изучались микросателлитные SSR (Simple Sequence Repeat) маркеры ДНК.
Помимо того, что маркеры должны обладать особой спецификой, они должны быть еще и хорошо различимыми белками. Поэтому заранее изучаются биологическая специфика, молекулярное строение и другие биохимические структуры данных белков. Так как гены располагаются в различных локусах хромосом и в целом составляют геном, основываясь на принцип ген-белок, используя их как маркер, можно получить информацию о генотипе [2,14].
Хорошо известна значимость белков глиадина и глютенина в хлебопечении [12]. Качество хлеба напрямую связано с наличием или отсутствием специальных белковых единиц [8]. Кроме того, качество хлеба зависит от соотношения мономерных белков к полимерным белкам, а также от качества и объема распространения полимерных белков [9]. Глютеновые белки, глиадины и глютенины составляют 80-85% от общих белков муки и тем самым они придают эластичность и растяжимость пшеничной муке [15].
Одним из актуальнейших вопрос целевой науки является изучение синтеза генетического надзора над запасными белками глиадин и глютенин, составляющими основу эндосперма. Так как количество клейковины в зерне составляет 80-85% от запасных белков, эти белки называют белками клейковины [3,7].
Материал и методика
В качестве материала исследований использовались местные сорта мягкой пшеницы Азербайджанской Республики Акинчи-84, Гобустан, Нурлу-99, Азаметли 95, Рузи-84, Гырмызыгюль, Баяз, Парзиван-1, Парзиван-2, Шеки-1, Шафаг, Дурдана, Бирлик, Достлуг, Бол-Бугда, интродуцированные Дагдаш, Анза, Безостая-1, Саратовская-29. В зерне эндосперма этих растений были изучены запасные белки и составлены электрофореограммы глиадинкодирующих локусов Gld 1A, Gld 1B, Gld 1D, Gld 6A, Gld 6B, Gld 6D. Электрофоретический анализ проводился по методу W.Bushuk, R.R.Zillman, модифицированный Ф.А.Попереля и др. [4,5].
Результаты и обсуждения
В проводимых нами научных исследованиях наличие в генах глиадинкодирующих локусов большого количества аллелей привело к расширению каталога компонентов блоков глиадин аллелей.
Идентификация исследуемых сортов согласно генетической формуле электрофореограмм была претворена в жизнь на основе глиадинкодирующих локусов Gld 1A, Gld 1B, Gld 1D, Gld 6A, Gld 6B, Gld 6D и Gld 2-1А.
Во время проведения исследований запасные белки глиадина были условно разделены на 4 зоны и получили название ω-, γ-, β-, α-глиадины. Глиадины, находящиеся в зерне пшеницы-гетерогенные белки. Глиадины, будучи в форме мономерной цепи, составляют 50% зерна и по молекулярному весу в полиакриламидном геле имеет 4 различные формы (α-глиадин, β-глиадин, γ-глиадин, ω-глиадин) [16]. Все глиадины считаются мономерами, несмотря на то, что у них могут быть дисульфидные связи (ω-глиадин), а также цепочковидные дисульфидные связи (α-, β-, γ-глиадины) [10,11].
Была проведена идентификация компонентов блоков глиадиналлелей двух местных сортов: Экинчи-84, Эземетли-95 на основе электрофореограмм глиадинкодирующих локусов (рис. 1).
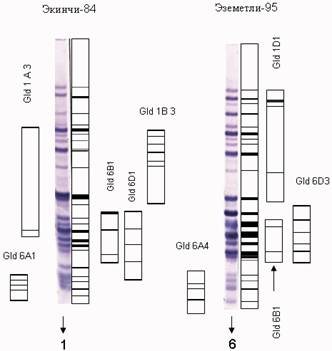
Рис. 1. Блоки компонентов аллелей в сортах мягкой пшеницы
Была определена частота встречаемости блоков компонентов аллелей в местных сортах пшеницы, таких как Акинчи-84, Gld 1A3, Gld 1B3, Gld 1D1, Gld 6A1, Gld 6B1, Gld 6D1 и Gld 2-1А3; блоков компонентов аллелей Гобустан, Gld 1A5, Gld 1B? новый, Gld 1D1, Gld 6A4, Gld 6B2, Gld 6D3 и Gld 2-1А3; Ниглу-99, Gld 1A5, Gld 1B3, Gld 1D1, Gld 6A4, Gld 6B2, Gld 6D2 и Gld 2-1А3; а также в сорте Гырмызыгюль, Gld 1A5, Gld 1B1, Gld 1D1, Gld 6A2, Gld 6B2, Gld 6D2. Однако, в сорте Гырмызыгюль Gld 2-1А3 блоков компонентов аллелей не встречалось.
Во время проведения электрофоретического анализа стало известно, что в зернах сортов мягкой пшеницы Гобустан, Нурлу-99, Рузи-84, Гырмызыгюль, Дурдана были идентифицированы глиадинкодирующие локусы, отвечающие за синтез запасных белков Gld 1A5, в сорте Азаметли-95, Gld 1A6, в сортах Парзиван-1, Шеки-1, Достлуг, Бирлик, Дагдаш, Безостая-1 (St) и Анза Gld A4; в сортах Акинчи-84, Бол-бугда, Баяз Gld 1A10, а в сорте Шафаг Gld 1A2. Таким образом, в местных сортах мягкой пшеницы частота повторяемости блоков компонентов глиадин аллелей различна. В этих сортах блоки компонентов аллелей Gld 1A5 по сравнению с блоками Gld 1A4 и Gld 1A6 встречаются довольно чаще. Блок Gld 1A5 в азербайджанских сортах встречается чаще, поэтому высокий показатель качества семян этих сортов соответственно выше. Это дает основание сделать вывод, что качество этих семян есть молекулярный белковый маркер.
В результате вышеизложенного можно заключить, что частота встречаемости глиадинкодирующих локусов аллелей в мягкой пшенице связана с генетическим аппаратом, этих же глиадинкодирующих локусов.
В вышеуказанных сортах Экинчи-84, Нурлу-99, Эземетли-95 и Дурдана Gld 1B3, Гырмызыгул, Рузи-84, Безостая-1, Анза, Бирлик, Достлуг, Перзиван-2, Шефег, Дагдаш Gld 1B1 была проведена идентификация блоков компонентов. В сортах мягкой пшеницы Экинчи-84, Саратовская-29, Бол-бугда, Беяз, парзиван-1 Gld 6D1, Нурлу-99, Гырмызыгуль, Анза, Дурдана Gld 6D2, Гобустан, Эземетли-95, Рузи-84, Дагдаш, Бирлик, Достлуг, Перзиван-2, Шеки-1 Gld 6D3, Безостая-1 (St), Шефег Gld 6D4 была проведена идентификация блоков компонентов аллелей.
Gld 1A4 блок компонентов наблюдался у классических сортов Безостая-1 (St), Дагдаш (Турция), Парзиван-2, Шеки-1. На основании литературных данных этот блок является маркерам качестве хлеба [6].
Наряду с хромосомой Gld 1A глиадинкодирующих локусов выявлен локус Gld 2-1A, который локализуется там же и контролирует синтез этого спектра.
В сортах Экинчи-84, Гобустан, Нурлу-99, Эземетли-95, Рузи-84, Бол-бугда, Бейаз, Парзиван-1 Gld 2-1A1 были выявлены блоки компонентов аллелей (табл. 1).
Во время исследований местных сортов (Гобустан, Бол-бугда, Беяз и Шеки-1) были обнаружены новые блоки компонентов аллелей (рис. 2, 3).
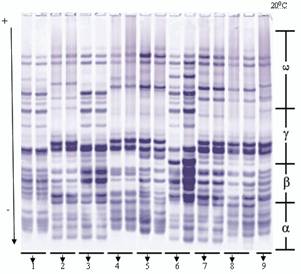
Рис. 2. Электрофореграммы глиадинокодирующих локусов мягкой пшеницы
1 - Экинчи-84, 2 - Гобустан, 3 - Нурлу-99, 4 - Гырмызыгул, 5 - Безостая (st),
6 - Эземетли- 95, 7- Анза, 8 - Рузи- 84, 9 - Саратовская-29
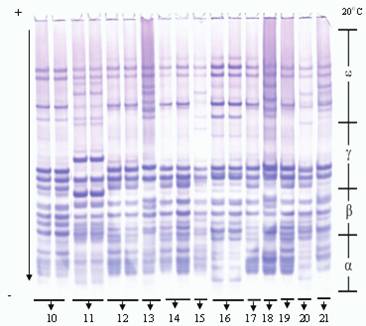
Рис. 3. Электрофореграммы запасных белков глиадина в сортах мягкой пшеницы
10 - Дагдаш, 11 - Дурдана, 12 - Бирлик, 13 - Бол-бугда, 14 - Безостая (st), 15 - Бейаз,
16 - Анза (marker), 17 - Достлуг, 18 - Парзиван-1, 19 - Парзиван-2, 20 - Шеки-1, 21 - Шефег
Таблица 1
Формулы глиадиновых локусов
|
s/s
|
Названия сорта |
Gld 1A |
Gld 1B |
Gld1D |
Gld 6A |
Gld 6B |
Gld 6D |
Gld 2-1A |
|
1 |
Экинчи-84 |
10 или 3 |
3 |
1 |
1 |
1 |
1 |
3 |
|
2 |
Гобустан |
5 |
новый |
1 |
4 |
2 |
3 |
3 |
|
3 |
Нурлу-99 |
5 |
3 |
1 |
4 |
2 |
2 |
3 |
|
4 |
Гырмызыгул |
5 |
1 |
1 |
2 |
2 |
2 |
- |
|
5 |
Безостая (st) |
4 |
1 |
1 |
2 |
1 |
4 |
- |
|
6 |
Эземетли- 95 |
6 |
3 |
1 |
4 |
1 |
3 |
3 |
|
7 |
Анза |
4 |
1 |
1 |
1 |
2 |
2 |
3 |
|
8 |
Рузи- 84 |
5 |
1 |
1 |
1 |
3 |
3 |
3 |
|
9 |
Саратовская-29 |
5 |
4 |
1 |
1 |
1 |
1 |
3 |
|
10 |
Дагдаш |
4 |
1 |
1 |
1 |
1 |
3 |
3 |
|
11 |
Дурдана |
5 |
3 |
1 |
3 |
1 |
2 |
1 |
|
12 |
Бирлик |
4 |
1 |
1 |
1 |
1 |
3 |
1 |
|
13 |
Бол-бугда |
10 или 3 |
новый |
1 |
1 |
1 |
1 |
3 |
|
14 |
Бейаз |
10 |
новый |
1 |
1 |
? |
1 |
3 |
|
15 |
Достлуг |
4 |
1 |
1 |
1 |
1 |
3 |
1 |
|
16 |
Парзиван-1 |
? |
4? |
1 |
2 |
1 |
1 |
3 |
|
17 |
Парзиван-2 |
4 |
1 |
1 |
1 |
1 |
3 |
1 |
|
18 |
Шеки-1 |
4 |
новый |
1 |
4 |
1 |
3 |
? |
|
19 |
Шефег |
2? |
1? |
1 |
1? |
1 |
4 |
1 |
Используя известные электрофоретические спектры сорта мягкой пшеницы «Анза» (Америка), а так же электрофореограммы электрофоретических спектров местных и других сортов мягкой пшеницы при помощи статистической программы «Био Капт» были подсчитаны молекулярная масса и другие показатели (объем спектра, высота, площадь и др.), (рис. 4).
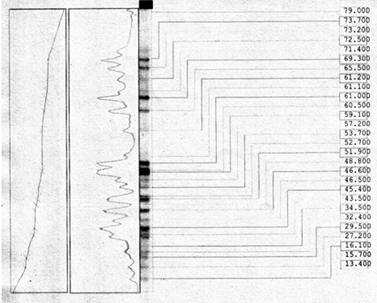
Рис. 4. Молекулярная масса у сорта мягкой пшеницы Анза
Объем 8 спектра «Анза» 348562, площадь 1564, молекулярная масса 64.200, 10 спектра объем 2770, площадь 23, молекулярная масса 61.500. Однако у местного сорта Экинчи-84 объем 1 спектра 431870, площадь 1794, молекулярная масса 97394 больше чем у «Анза» (маркер). Был подсчитан объем 10 спектра 2990, площадь 23, молекулярная масса 59.787. Объем 7 электрофоретического спектра Гобустан 398312, площадь 1728, молекулярная масса 64.690, объем 13 спектра 4272, площадь 24, молекулярная масса 52.498, У Гырмызыгюль объем 16 электрофоретического спектра 514028, при площади 2300, молекулярная масса 34.116, что отличает его от «Анза» и других сортов высокими показателями. Объем 14 спектра 3012, площадь 23, молекулярная масса 44281. В результате анализов молекулярная масса запасных белков глиадина и высота спектра резко отличалась по сортам. Однако объём электрофоретических спектров и их площадь в значительной степени отличалась от сорта «Анза», взятого как маркер (табл. 2).
Таблица 2
Электрофоретические спектральные анализы глиадина в сортах Анза, Экинчи-84, Гобустан и Гырмызыгуль мягкой пшеницы
|
Анза |
Объем |
высота |
площадь |
молекулярная масса |
Экинчи-84 |
объем |
высота |
Площадь |
молекулярная масса |
|
1спектр |
67822 |
250 |
360 |
79.000 |
1 спектр |
431870 |
255 |
1794 |
97.394 |
|
2спектр |
3170 |
246 |
23 |
73.700 |
2 спектр |
59540 |
250 |
276 |
73.700 |
|
3спектр |
48218 |
248 |
276 |
73.200 |
3 спектр |
4304 |
244 |
23 |
73.200 |
|
4спектр |
79524 |
250 |
414 |
72.500 |
4 спектр |
286782 |
254 |
1288 |
72.191 |
|
5спектр |
121064 |
250 |
621 |
71.400 |
5 спектр |
3666 |
242 |
23 |
67.424 |
|
6спектр |
2834 |
238 |
23 |
69.300 |
6 спектр |
78986 |
252 |
391 |
65.734 |
|
7спектр |
97248 |
252 |
529 |
65.500 |
7 спектр |
68040 |
252 |
322 |
65.201 |
|
8спектр |
348562 |
252 |
1564 |
64.200 |
8 спектр |
3746 |
244 |
23 |
64.526 |
|
9спектр |
54632 |
246 |
299 |
62.900 |
9 спектр |
371538 |
254 |
1702 |
63.947 |
|
10спектр |
2770 |
228 |
23 |
61.500 |
10 спектр |
2990 |
244 |
23 |
59.787 |
|
11спектр |
40470 |
240 |
299 |
60.300 |
11 спектр |
24390 |
250 |
161 |
58.629 |
|
12спектр |
5626 |
236 |
46 |
59.100 |
12 спектр |
145974 |
252 |
713 |
53.141 |
|
13спектр |
43606 |
246 |
253 |
57.200 |
13 спектр |
74190 |
254 |
414 |
51.013 |
|
14спектр |
115350 |
248 |
575 |
53.700 |
14 спектр |
3264 |
244 |
23 |
49.626 |
|
15 спектр |
10164 |
232 |
69 |
52.700 |
15 спектр |
66110 |
252 |
391 |
47.028 |
|
16 спектр |
71030 |
246 |
391 |
51.900 |
16 спектр |
32750 |
252 |
184 |
46.317 |
|
17 спектр |
2968 |
232 |
23 |
48.800 |
17 спектр |
25278 |
252 |
138 |
45.717 |
|
18 zolaq |
106968 |
244 |
621 |
46.600 |
18 спектр |
208558 |
255 |
989 |
36.883 |
|
19 спектр |
2956 |
222 |
23 |
45.500 |
19 спектр |
8362 |
252 |
46 |
32.211 |
|
20 спектр |
41310 |
240 |
276 |
43.500 |
20 спектр |
16748 |
252 |
92 |
31.468 |
|
21 спектр |
100802 |
248 |
529 |
34.500 |
21 спектр |
56258 |
254 |
299 |
29.364 |
|
22 спектр |
30654 |
246 |
161 |
32.400 |
|
|
|
|
|
|
23 спектр |
75222 |
246 |
368 |
29.500 |
|
|
|
|
|
|
24 спектр |
105852 |
248 |
506 |
27.200 |
|
|
|
|
|
|
25 спектр |
91473 |
240 |
470 |
16.100 |
|
|
|
|
|
|
26 спектр |
97651 |
235 |
450 |
15.700 |
|
|
|
|
|
|
27 спектр |
8965 |
230 |
440 |
13.400 |
|
|
|
|
|
|
Гобустан |
объем |
высота |
пло-щадь |
молекулярная масса |
Гырмыз-ыгуль |
объем |
высота |
пло-щадь |
молекулярная масса |
|
1 спектр |
345038 |
252 |
1488 |
97.706 |
1 спектр |
390020 |
246 |
1748 |
79.312 |
|
2 спектр |
69590 |
246 |
336 |
79.000 |
2 спектр |
8238 |
224 |
46 |
74.051 |
|
3 спектр |
56618 |
246 |
288 |
74.051 |
3 спектр |
45250 |
240 |
230 |
73.399 |
|
4 спектр |
8628 |
238 |
48 |
73.200 |
4 спектр |
217430 |
240 |
1081 |
72.254 |
|
5 спектр |
204092 |
248 |
960 |
70 70.974 |
5 спектр |
7416 |
208 |
46 |
69 69.505 |
|
6 спектр |
140764 |
250 |
648 |
65.994 |
6 спектр |
514028 |
248 |
2300 |
63.330 |
|
7 спектр |
398312 |
250 |
1728 |
64.690 |
7 спектр |
2642 |
158 |
23 |
62.041 |
|
8 спектр |
56012 |
238 |
312 |
62.900 |
8 спектр |
39722 |
206 |
299 |
60.815 |
|
9 спектр |
2744 |
192 |
24 |
61.500 |
9 спектр |
216960 |
246 |
1081 |
53.347 |
|
10 спектр |
31376 |
218 |
240 |
60.496 |
10 спектр |
3792 |
206 |
23 |
52.098 |
|
11 спектр |
64908 |
250 |
360 |
57.734 |
11 спектр |
54316 |
240 |
276 |
50.138 |
|
12 спектр |
151142 |
250 |
672 |
54.187 |
12 спектр |
118354 |
242 |
552 |
49.291 |
|
13 спектр |
4272 |
240 |
24 |
52.498 |
13 спектр |
57488 |
216 |
345 |
46.243 |
|
14 спектр |
23176 |
244 |
120 |
51.544 |
14 спектр |
3012 |
164 |
23 |
44.281 |
|
15 спектр |
48400 |
246 |
216 |
51.013 |
15 спектр |
62230 |
216 |
368 |
37.292 |
|
16 спектр |
8270 |
238 |
48 |
49.626 |
16 спектр |
52210 |
212 |
322 |
34.116 |
|
17 спектр |
113318 |
244 |
600 |
47.706 |
17 спектр |
3146 |
180 |
23 |
32.589 |
|
18 спектр |
6156 |
226 |
48 |
46.000 |
18 спектр |
52110 |
224 |
322 |
30.095 |
|
19 спектр |
39108 |
238 |
264 |
44.488 |
|
|
|
|
|
|
20 спектр |
144858 |
246 |
720 |
34.308 |
|
|
|
|
|
|
21 спектр |
4596 |
242 |
24 |
32.779 |
|
|
|
|
|
|
22 спектр |
68264 |
246 |
336 |
30.418 |
|
|
|
|
|
|
23 спектр |
103502 |
250 |
504 |
27.734 |
|
|
|
|
|
После проведения глиадиновой экстракции и электрофореза в зёрнах мягкой пшеницы, генетической индекс схожести между сортами был пронумерован по методу «1» и «0». Колонки, стоящие в одном и том же месте были пронумерованы «1», другие же неподходящий по площади колонки-пронумерованы «0», индекс схожести был подсчитан на основании статистической программы SPSS. Для того, чтобы выявить генетическую близость образцов по индексу схожести используется кластерный анализ и составляется дендрограмма. При подсчёте индекса генетической схожести стало известно, что у сортов Нурлу-99, Дагдаш, Рузи-84, и Эземетли-95, Шеки-1 индекс схожести между ними более близок, у сортов Гырмызыгуль, Саратовская-29- сравнительно близок. Вместе с тем, между сортами мягкой пшеницы Гобустан, Рузи-84, Саратовская-29, Дурдана, Парзиван-1, Дагдаш и Шеки-1 индекс схожести был довольно далек (табл. 3).
Таблица 3
Индексы схожести между сортами мягкой пшеницы
|
SСорта |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
|
1. Экинчи-84 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2. Гобустан |
,375 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3. Нурлу-99 |
,625 |
,364 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4. Гырмызыгуль |
,310 |
,257 |
,300 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5. Эземетли-95 |
,464 |
,382 |
,448 |
,323 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
6. Рузи-84 |
,577 |
,343 |
,909 |
,323 |
,419 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
7. Саратовская-29 |
,407 |
,333 |
,345 |
,652 |
,323 |
,367 |
1 |
|
|
|
|
|
|
|
|
|
|
|
8. Дагдаш |
,625 |
,324 |
,905 |
,300 |
,400 |
,909 |
,345 |
1 |
|
|
|
|
|
|
|
|
|
|
9. Дурдана |
,444 |
,286 |
,600 |
,345 |
,500 |
,556 |
,393 |
,538 |
1 |
|
|
|
|
|
|
|
|
|
10. Бирлик |
,500 |
,455 |
,433 |
,313 |
,500 |
,406 |
,500 |
,387 |
,483 |
1 |
|
|
|
|
|
|
|
|
11. Бол-бугда |
,303 |
,361 |
,222 |
,265 |
,394 |
,243 |
,265 |
,257 |
,222 |
,306 |
1 |
|
|
|
|
|
|
|
12. Баяз |
,345 |
,324 |
,250 |
,393 |
,500 |
,235 |
,444 |
,250 |
,379 |
,344 |
,222 |
1 |
|
|
|
|
|
|
13. Достлуг |
,314 |
,405 |
,306 |
,353 |
,441 |
,324 |
,353 |
,306 |
,469 |
,351 |
,457 |
,424 |
1 |
|
|
|
|
|
14. Парзиван-1 |
,462 |
,375 |
,444 |
,310 |
,577 |
,414 |
,407 |
,444 |
,444 |
,500 |
,303 |
,393 |
,314 |
1 |
|
|
|
|
15. Парзиван-2 |
,270 |
,395 |
,231 |
,270 |
,389 |
,250 |
,237 |
,231 |
,263 |
,378 |
,529 |
,231 |
,528 |
,205 |
1 |
|
|
|
16. Шеки-1 |
,500 |
,412 |
,483 |
,313 |
,667 |
,552 |
,400 |
,483 |
,433 |
,484 |
,343 |
,433 |
,389 |
,500 |
,308 |
1 |
|
|
17. Шефег |
,345 |
,324 |
,333 |
,393 |
,400 |
,313 |
,393 |
,333 |
,333 |
,303 |
,189 |
,481 |
,237 |
,393 |
,200 |
,387 |
1 |
У сортов мягкой пшеницы при помощи мономерных проломиновых белковых маркеров была исследована генетическая близость и, используя статистическую программу NTSYS-pc (numerical taxonomy and multivarion analysis sistem-2.00) была построена дендрограмма. Как видно на рисунка 5 дендрограмма делится на основную большую и две малые части. Первая большая часть делится на два яруса. В первом ярусе сгруппировались сорта Экинчи-84, Нурлу-99, Рузи-84, Дагдаш, Дурдана а во втором ярусе сорта Эзаматли-95, Шеки-1, Парзиван-1, Шнфег и Бирлик, генетически более близкие друг другу. Сорта Гырмызыгуль, Саратовская-29, и Баяз составили вторую малую часть, а сорта Гобустан, Бол-бугда, Достлуг и Парзиван-2 сгруппировались в третьей малой части (рис. 5) [13].
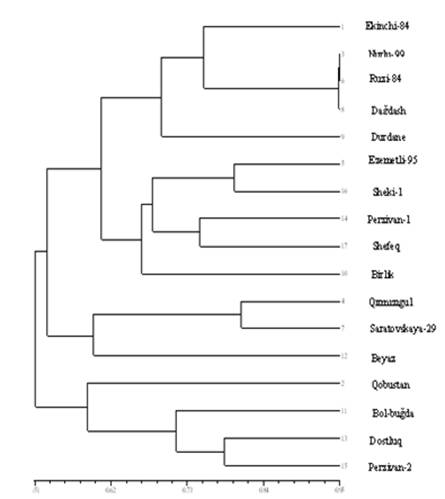
Рис. 5. Группировка сортов мягкой пшеницы по глиадиновым маркерам
СПИСОК ЛИТЕРАТУРЫ
- 1. Mahmudov R.U., Buğda dəninin qliadin zülalları və seleksiyada ondan istifadə imkanları. Az.Elmi-Tədqiqat Əkinçilik İnstitutunun elmi əsərləri məcmuəsi XXI cild. Bakı-2005, s.191-196
- 2. Конарев А.В., Конарев В.Г.,
Губарева Н.К., Пенева Т.И. Белки семян как маркеры в решении проблем генетических ресурсов растений, селекции и семеноводства. Цитология и генетика, 2000, т. 34, № 2, с. 91-104. - 3. Попереля Ф.А., Бито М., Созинов А.А. Связь блоков компонентов глиадина с выживаемостью растений и их продуктивностью, окраской колоса и качеством гибридов F2 от скрещивания сортов Безостая 1 и Црвена Звезда // Докл. ВАСХНИЛ. - 1980. № 4. С. 4-7.
- 4. Попереля Ф.А. и др. Определение гибридности семян кукурузы по электрофоретическим спектрам зеина. Доклады ВАСХНИЛ, 1989, № 3.
- 5. Созинов А.А., Попереля Ф.А. Методика вертикального дискового електрофореза белков в крахмальном геле.// Информационный бюллетень СЕВ, 1974, вып.№1, с.135-144.
- 6. Созинов А.А. Полиморфизм белков и его значение в генетики и селекции. Москва, «Наука», 1985, с. 272.
- 7. Bashuk W. and Zillman R.R. Wheat cultivar identification by gliadin electrophore-grams. I.Apparatus, method and nomenclature. Cand. J.Plant Sci., 1978, vol. 58, p. 505-515.
- 8. Gupta. R.B., Singh. N.K., Shepherd. K.W. 1989. The cumulative effect of allelic variation in LMW and HMW glutenin subunits on dough properties in the progeny of two bread wheats. Theoretical and Applied Genetics 77, 57-64.
- 9. Johansson. E., Prieto-Linde. M.L. and Jönsson. J.Ö. 2001. Effects of wheat cultivar and nitrogen application on storage protein composition and breadmaking quality. Cereal Chemistry 78, 19-25.
- 10. Müller. S., Wieser. H. 1995. Disulphide bonds of alfa-type gliadins. Journal of Cereal Science 22, 21-27.
- 11. Müller. S., Wieser. H. 1997. The location of disulphide bonds in monomeric gama-gliadins. Journal of Cereal Science 26, 169-176. 36.
- 12. Morrison. W.R. 1988. Lipids. In Wheat: chemistry and technology, Pomeranz, Y., ed. (St.Paul., USA, American Association of Cereal Chemists), 373-439 pp.
- 13. Rohlf F.J. (1998) NTSYS-PC. (Numerical Taxonomy and Multivarion Analysis Sistem, version 1.80) Exeter Software, Setauket, NY.
- 14. Shepherd K.W. Chromosomal control of endosperm proteins in wheat and rye // Proc. 3rd Intern. Wheat Genet. Sump., Canberra, Austral. Acad. Sci., 1968, p. 86-96.
- 15. Shewry P.R., Napier J.A., Tatham A.S. 1995. Seed storage proteins: structures and biosynthesis. The Plant Cell 7, p.945-956.
- 16. Woychik J. H., Boundy J. A., Dim-
ler R.J. 1961. Starch gel electrophoresis of wheat gluten proteins with concentrated urea. Archieval Biochemistry and Biophysics 94, p.477-482.
Библиографическая ссылка
Каримов А.Я ИССЛЕДОВАНИЕ ИНДЕКСА ГЕНЕТИЧЕСКОЙ СХОЖЕСТИ И ИДЕНТИФИКАЦИЯ СОРТОВ МЯГКОЙ ПШЕНИЦЫ СПОСОБОМ ГЛИАДИНОВОГО МАРКЕРА // Современные проблемы науки и образования. 2009. № 6-3. ;URL: https://science-education.ru/ru/article/view?id=1421 (дата обращения: 01.07.2026).