



Введение
Каждую особь любого вида живых организмов характеризует возраст. Различают календарный возраст, измеряемый физическими единицами времени, и биологический возраст, определяемый по совокупности различных морфологических, физиологических, биохимических и т.п. признаков организма. Использование понятия биологический возраст оказывается необходимым в популяционной биологии растений и лишайников, особенно в тех случаях, когда определение календарного возраста не представляется возможным [7]. В индивидуальном развитии (онтогенезе) особи растения и лишайника на основе совокупности морфологических маркерных признаков различают последовательные онтогенетические периоды – латентный, прегенеративный, генеративный и постгенеративный. В пределах каждого из них определяют последовательные дискретные онтогенетические состояния.
Таким образом, непрерывный процесс онтогенеза растений и лишайников описывается набором последовательных онтогенетических состояний, и онтогенетическое состояние является качественным (не количественным!) признаком особи. Заметим, что поскольку последовательные онтогенетические состояния упорядочены во времени, календарный и биологический возраст особей неизбежно сильно скоррелированы, при этом особи одного онтогенетического состояния могут иметь разный календарный возраст.
А.А. Уранов [7], исходя из логистической кривой роста особи в течение онтогенеза, ввел весовой коэффициент (численную характеристику) каждого онтогенетического состояния. Рассматривая совокупность особей популяции, можно построить онтогенетический спектр популяции.
Методика сбора материала для характеристики онтогенетического спектра популяции заключается в том, что каждая выборка включает несколько субвыборок, собранных в разных частях популяции, в разные годы и т.п. При исследовании, например, эпифитных лишайников учитываются особи (слоевища) всех онтогенетических состояний отдельно на каждом дереве, с последующим суммированием данных в пределах местообитания или определенного форофита. Таким образом, единицей наблюдения здесь является отдельное дерево.
Однако суммирование материалов субвыборок (дерево для эпифитных лишайников) и рассмотрение данных по выборке в целом правомочно только в случае гомогенности выборки – распределения разных субвыборок в пределах выборки статистически значимо не различаются, т. е. они могут быть описаны одним полиномиальным распределением [10].
Цель настоящей работы – предложить методы статистического анализа онтогенетической структуры популяции эпифитного лишайника Pseudevernia furfuracea (L.) Zopf с учетом избыточной изменчивости, гетерогенности субвыборок в пределах выборки.
Материалы и методы
Слоевища P. furfuracea собирали с деревьев, произрастающих на 9 пробных площадях на территории Республики Марий Эл – табл. 1. Две пробные площади заложены на территории Государственного природного заповедника «Большая Кокшага». Одна – в сосняке зеленомошном, деревья сосны обыкновенной (Pinus sylvestris L.) представлены двумя классами возраста: бо́льшая часть (85 %) в возрасте 60–80 лет, выборка слоевищ 1a; остальные деревья (15 %) в возрасте 100–120 лет, выборка слоевищ 1b. Другая пробная площадь заложена также в сосняке зеленомошном с участием березы повислой (Betula pendula Roth.). Слоевища P. furfuracea собирали с рядом расположенных деревьев сосны обыкновенной (выборка 2a) и березы повислой (выборка 2b).
Три пробные площади заложены на территории Старожильского лесничества Медведевского района в окрестностях верхового болота: выборка 3 взята на деревьях сосны в сосняке черничном, примыкающем к болоту; выборка 4 – на сосне в сосняке кустарничково-сфагновом непосредственно на болоте; выборка 5 – на березе пушистой (Betula pubescens Ehrh.) в березняке осоково-белокрыльниково-сфагновом у противоположного края болота.
Четыре выборки взяты на сосне на территории Керебелякского лесничества Национального парка «Марий Чодра»: на рядом расположенных участках сосняка брусничного – выборка № 6 (в мезоповышении) и выборка № 7 (в мезопонижении); на рядом расположенных участках сосняка лишайниково-мшистого – выборка № 8 (в мезоповышении) и выборка № 9 (в мезопонижении). На каждом дереве слоевища учитывали до высоты 4 м, определяя их онтогенетические состояние, описанное в [4]. Выборка № 4 включает 47 деревьев, в остальных выборках – по 10–20 деревьев. Число слоевищ на дереве варьирует от нескольких до полутора сотен, только в выборке № 8 число слоевищ варьирует от 64 до 238 и в выборке № 9 – от 93 до 359.
В природных популяциях лишайников онтогенетические спектры могут быть построены, начиная с v1 онтогенетического состояния, поскольку слоевища начальных онтогенетических состояний имеют микроскопические размеры, а видовую принадлежность im особей иногда трудно идентифицировать.
Таблица 1. Характеристика местообитаний P. furfuracea
|
Местоположение |
№ пробной площади |
Тип леса, состав древостоя |
Бонитет древостоя |
Полнота |
№ выборки |
Форофит |
Характеристика древостоя |
||
|
Возраст, г |
Высота, м |
Диаметр ствола, см |
|||||||
|
Заповедник "Большая Кокшага" |
1 |
Сосняк зеленомошный, 10С+Б |
I |
0,7 |
1a |
сосна |
65 |
21 |
22 |
|
1b |
сосна |
110 |
26 |
43 |
|||||
|
2 |
Сосняк зеленомошный, 9С1Б |
II |
0,8 |
2a |
сосна |
65 |
20 |
20 |
|
|
2b |
береза повислая |
45 |
16 |
15 |
|||||
|
Старожильское лесничество |
3 |
Сосняк черничный, 10С |
II |
0,8 |
3 |
сосна |
70 |
21 |
16 |
|
4 |
Сосняк кустарничково-сфагновый, 10С |
V |
0,5 |
4 |
сосна |
110 |
15 |
20 |
|
|
5 |
Березняк осоково-белокрыльниково-сфагновый, 10Б |
II |
0,7 |
5 |
береза пушистая |
62 |
17 |
16 |
|
|
Керебелякское лесничество |
6 |
Сосняк брусничный, 7С3Б |
I |
0,8 |
6 |
сосна |
65 |
21 |
29 |
|
7 |
7 |
сосна |
65 |
24 |
28 |
||||
|
8 |
Сосняк лишайниково-мшистый, 6С4Б |
II |
0,7 |
8 |
сосна |
60 |
26 |
20 |
|
|
9 |
9 |
сосна |
60 |
27 |
20 |
||||
Для проверки однородности онтогенетических спектров субвыборок в пределах выборки использовали критерий хи-квадрат [10]. Для анализа онтогенетических спектров проводили оценку и сравнение параметров онтогенетических спектров. Использовались следующие параметры: индекс возрастности популяции Δ (средняя возрастность популяции) [7], индекс восстановления I1 (отношение числа прегенеративных особей к сумме прегенеративных и генеративных) [1]. Так как можно рассматривать параметр I1 как вероятность в биномиальном распределении, то проводилась оценка дисперсии данного параметра для выборки с учетом и без учета гетерогенности субвыборок в пределах выборки [2].
Значения параметров были вычислены для каждого дерева. Для оценки значения параметров выборок, учитывая гетерогенность субвыборок, использовалась процедура ресамплинга [8]: проводилась бутстреп-процедура для распределений субвыборк в пределах выборки, для полученной псевдо-выборки вычислялось значение индекса. Медиана распределения значений индекса выборки, полученных в ходе рандомизации, является точечной оценкой параметра, доверительный интервал для оценки находится из соответствующих квантилей. Для сравнения значений параметров проводили дисперсионный анализ (модель I) и множественные сравнения с помощью критерия Шеффе. Чтобы обойти условия применения обычного дисперсионного анализа [10], использовали рандомизационную процедуру [8]: осуществлялось случайное перемешивание значений параметра субвыборок между выборками без повторений, для данных полученных после перестановки вычислялось F-значение, в результате рандомизационной процедуры получали распределение F-значений. Доля значений, больших чем F-значение, полученное для исходных данных, есть статистическая значимость разницы между выборками.
Для оценки доли влияния изменчивости между онтогенетическими спектрами выборок в общей изменчивости проводили дисперсионный анализ (модель II), используя бутстреп (извлечение с повторением значений параметра для площадок в каждой выборке) [8]. Для оценки доли влияния использовали также регрессионную модель для упорядоченных классов [9]. Для того чтобы учесть объемы выборок и субвыборок, проводили процедуру взвешивания. Во всех процедурах ресамплинга число итераций равно 10000.
При анализе данных использовали среду «R» и пакет «Statistica», для сравнения параметров онтогенетических спектров была использована программа «OntoParam» [3], для построения регрессионной модели для упорядоченных классов был использован пакет «ordinal» среды «R».
Результаты и обсуждение
На рисунке 1 показаны онтогенетические спектры популяций (выборок), жирная линия – суммарные данные. Можно видеть, что все спектры выраженно положительно асимметричны (правый хвост распределения длиннее левого) с максимумом на особях v2 онтогенетического состояния. Заметим, что частота g3, ss и s особей в природных популяциях P. furfuracea очень мала (в среднем 0,4 %), поскольку они плохо удерживаются на стволе дерева [5, 6]. Это делает бессмысленным анализ индекса старения I2.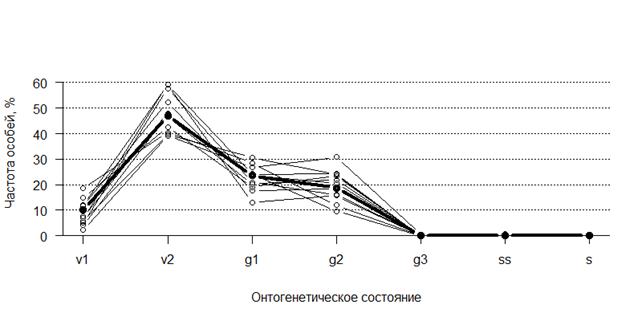
Рис. 1. Онтогенетические спектры выборок P. furfuracea, жирная линия – суммарный спектр
Изменчивость спектров разных выборок охватывает довольно широкую полосу, но возникает вопрос, можно ли их сравнивать, объединяя в каждой выборке данные по субвыборкам? Частоты v1 особей гетерогенны во всех изучаемых выборках, т.е. субвыборки различаются статистически значимо в пределах своей выборки (p = 0,0018-10-42), частоты особей других состояний гетерогенны в 6-7 выборках (p = 0,033-10-11). Отношение величины наблюдаемой дисперсии к ожидаемой биномиальной дисперсии (при случайных флуктуациях субвыборок) может достигать для частот разных онтогенетических состояний 10. Точно так же выявляется избыточная изменчивость между субвыборками по коэффициенту восстановления I1. Если пренебречь избыточной изменчивостью и попарно сравнивать онтогенетические спектры разных выборок, то из 55 попарных сравнений статистически значимыми на 1 %-ом уровне оказываются 50! Проводимый далее анализ учитывает эту избыточную изменчивость.
Результаты сравнения индексов возрастности Δ разных выборок показаны на рис. 2. Значения Δ для разных выборок распределены по шкале довольно равномерно. На 1 %-м уровне значимы лишь 3 парных разности, на 5 % - 6 разностей из возможных 55. Как было отмечено выше, сравнение онтогенетических спектров без учета изменчивости между субвыборками в пределах выборки выявляют 50 значимых разностей из 55, при этом на гораздо более высоком уровне значимости. Различия между величинами Δ могут быть связаны с особенностями деревостоев, с особенностями субстратов. Так, на 5 %-м уровне различаются выборки с близко расположенных пробных площадей, но на разных форофитах - выборки 3 и 5; выборки 8 и 9 на сосне, в одном типе леса, но при разном положении в рельефе (повышение и понижение). В то же время ничем не выделяется выборка 4 в сосняке кустарничково-сфагновом на верховом болоте.
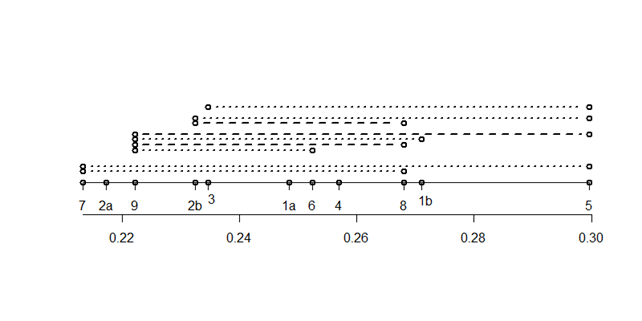
Рис. 2. Сравнение индексов возрастности Δ разных выборок. Внизу – шкала значений Δ; выше положение выборок на шкале Δ; - - - p < 0,01; ××××× p < 0,01
Индекс восстановления I1, тесно скоррелирован с индексом возрастности Δ (коэффициент ранговой корреляции Спирмена равен -0,95, p<10-4), что связано с очень малой долей g3, ss и s особей. По I1 выявляются статистически значимые различия на 5 %-м уровне только для 5 пар выборок (рис. 3).
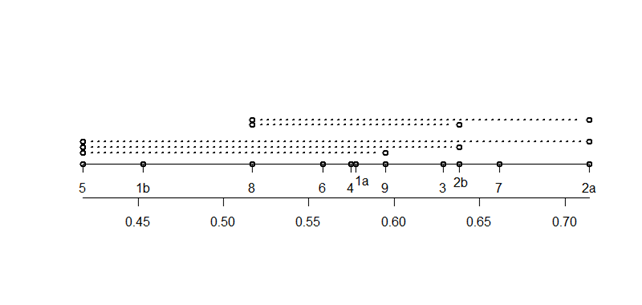
Рис. 3. Сравнение индексов восстановления I1 разных выборок. Внизу – шкала значений I1; выше положение выборок на шкале I1; ××××× p < 0,01
Оценки доли влияния межвыборочной изменчивости в общей изменчивости, проведенные с помощью ресамплинга на основе модели II рандомизационного дисперсионного анализа, дали для Δ 0,339 (95 %-ый доверительный интервал: 0,181–0,514) и для I1 – 0,204 (0,087-0,351). Наглядное представление о сравнительной величине меж- и внутригрупповой изменчивости онтогенетических спектров дают результаты применения регрессии для упорядоченных классов (рис. 4). Здесь выборки упорядочены по возрастанию значений средних для выборки эффектов (темные кружки), и в пределах каждой выборки видна большая изменчивость онтогенетических спектров P. furfuracea на отдельных деревьях в пределах выборки (светлые кружки). Количественная оценка доли межвыборочной изменчивости дает 0,297.
Таким образом, все три подхода (анализ Δ, I1 и регрессия для упорядоченных классов) дают сходные результаты: основная изменчивость онтогенетических спектров P. furfuracea, несмотря на разнообразие экологических условий и субстратов разных выборок, сосредоточена внутри выборок (между спектрами разных деревьев на одной пробной площади). В значительной мере это определяется, скорее всего, небольшим числом градаций онтогенетических состояний в спектрах выборок P. furfuracea.

Рис. 4. Результаты применения регрессии для упорядоченных классов к анализу изменчивости онтогенетических спектров выборок P. furfuracea. Объяснения в тексте
Выводы
- Онтогенетические спектры популяций эпифитного лишайника P. furfuracea выраженно левовершинные с максимумом на v2 онтогенетическом состоянии. Слоевища онтогенетических состояний, начиная с g3, практически отсутствуют (их доля не превышает 0,4 %).
- Онтогенетические спектры субвыборки в пределах выборки сильно различаются. Об этом свидетельствует варьирование частот отдельных онтогенетических состояний, и систематическое превышение наблюдаемой дисперсии частот онтогенетических состояний по отношению к биномиальной (до 10 раз).
- Результаты анализа без учета внутривыборочных различий онтогенетических спектров сильно расходятся с результатами анализа с учетом этих различий, приводя к ложным выводам.
- Доля межвыборочной изменчивости в общей изменчивости онтогенетических спектров составляет: для Δ – 0,339, для I1 – 0,204, для регрессии упорядоченных классов – 0,297. Таким образом, применение всех методов анализа указывает на то, что бо́льшая изменчивость онтогенетических спектров сосредоточена в пределах выборок.
Настоящая работа выполнена при поддержке Гранта РФФИ (проект № 12-04-01251).
Рецензенты:
Безель В.С., д.б.н., профессор, в.н.с., Института экологии растений и животных УрО РАН, г. Екатеринбург.
Воскресенская О.Л., д.б.н., профессор, зав. кафедрой экологии ФГБОУ ВПО «Марийский государственный университет» Министерства образования и науки РФ, г. Йошкар-Ола.
Библиографическая ссылка
Глотов Н.В., Софронов Г.Ю., Иванов С.М., Теплых А.А., Суетина Ю.Г. ОНТОГЕНЕТИЧЕСКИЕ СПЕКТРЫ ПОПУЛЯЦИЙ ЭПИФИТНОГО ЛИШАЙНИКА PSEUDEVERNIA FURFURACEA (L. ) ZOPF // Современные проблемы науки и образования. 2014. № 3. ;URL: https://science-education.ru/ru/article/view?id=13744 (дата обращения: 30.06.2026).