



Половая структура популяции представляет собой количественное соотношение самцов и самок в разных возрастных группах. Различают первичное, вторичное и третичное соотношение полов: первичное отражает соотношение самцов и самок в момент формирования зиготы, вторичное – среди новорожденных особей, третичное – в момент наступления половой зрелости [6]. Амфиподы являются раздельнополыми животными, и, как у подавляющего большинства таких организмов, их пол определяется на генотипическом уровне [13]. Как известно, такой механизм обеспечивает первичное соотношение полов равным 1:1. Однако в популяциях амфипод часто наблюдаются отклонения от этого классического соотношения. Известно о существовании некоторых внешних и внутренних факторов, которые могут оказывать влияние на процесс формирования пола у этих беспозвоночных. К таким факторам относятся температура [12], фотопериод [9, 15] и присутствие паразитов [9]. Отклонения в соотношении полов могут также появиться в результате разной продолжительности жизни [11, 16] или скорости созревания самцов и самок [10], в результате пространственной сегрегации самцов и самок по горизонтам литорали [8, 7, 17] или в результате различной смертности самцов и самок на ювенильных стадиях [11, 14].
В настоящей статье анализируется половая структура популяции Gammarus oceanicus – амфипод, являющихся одними из самых массовых на побережье северных морей. Бокоплавы этого вида являются полицикличными животными и размножаются несколько раз на протяжении жизни [5]. Период размножения G. oceanicus в Кольском заливе начинается в ноябре, когда появляются первые копулирующие пары, и заканчивается в августе. В сентябре и октябре наблюдается перерыв в размножении, характерный для всех популяций данного вида. Самки откладывают первую порцию яиц в выводковые сумки в декабре и вынашивают их на протяжении всех зимних месяцев. В марте из этих яиц появляется первая генерация молоди. После этого самки линяют и откладывают очередную порцию яиц. Всего за период размножения в популяции из Кольского залива отмечается появление четырех генераций – в марте, начале мая, середине июня и в августе [2]. Нашей задачей было проследить сезонную динамику соотношения полов в рассматриваемой популяции G. oceanicus и выяснить, оказывает влияние температура и фотопериод на формирование пола у особей этого вида.
Материал и методика
Для выполнения поставленной задачи были проанализированы 42 бентосные пробы, собранные на литорали в южном колене Кольского залива в районе Абрам-мыса в период с февраля 2002 г. по январь 2003 г. (рис. 1). Отбор проб производился ежемесячно, в периоды максимальных отливов. При взятии всех проб использовалась рамка размером 0,25 м2. Собранный материал фиксировали 4% формалином. В камеральных условиях устанавливали видовую принадлежность гаммарусов [5] и определяли пол. Самцов идентифицировали по наличию у них семенных мешков на вентральной части последнего переонального сегмента, самок – по наличию элементов марсупиальных сумок в основании первых четырех пар переопод. Проверка статистических гипотез осуществлялась с помощью метода χ2.
Рис. 1. Карта-схема расположения района исследований
Результаты и обсуждение
Анализ сезонной динамики соотношения полов в поселении
G. oceanicus в районе Абрам-мыса в 2002-2003 гг. показал, что на протяжении большей части года соотношение самцов и самок близко к 1:1 с небольшим преобладанием самок (рис. 2). Статистически значимое отклонение от классического соотношения наблюдалось лишь в отдельные месяцы. В конце февраля, августе и сентябре в поселении преобладали самцы, причем в сентябре их численность превышала численность самок в 2,8 раза (p<0,01). В июне большинство составляли самки (p<0,05).
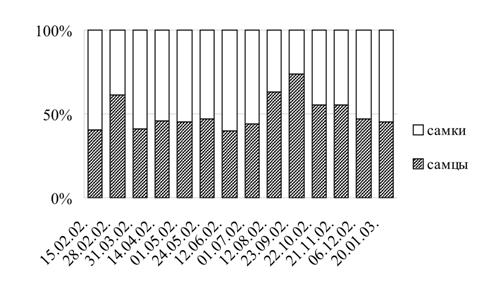
Рис. 2. Соотношение самцов и самок в поселении G. oceanicus в южной части Кольского залива (Абрам-мыс) в 2002-2003 гг.
Из литературных данных известно, что в других популяциях этого вида часто отмечаются неравное соотношение полов в течение года. Так, на побережье пролива Великая Салма Кандалакшского залива Белого моря в 1972 году летом преобладали самцы, осенью соотношение сдвигалось в сторону самок, зимой и ранней весной большинство составляли самки [1]. В губе Ярнышной Баренцева моря в период с 1978 по 1982 гг. летом всегда преобладали самцы, к началу перерыва в размножении соотношение полов выравнивалось, а зимой большинство составляли самки [3]. В двух бухтах Кандалакшского залива Белого моря в течение 1997 года соотношение полов изменялось по-разному. В бухте Круглой весной преобладали самки, летом – самцы, а осенью, к началу перерыва в размножении, соотношение полов выравнивалось. В бухте Иванов Наволок весной и осенью большинство составляли самки, а летом количество самцов и самок становилось примерно одинаковым [4].
Одной из причин сезонной динамики соотношения полов авторы вышеуказанных исследований называют преобладание в каждом поколении гаммарусов особей одного пола, что может быть связано с температурными условиями в момент формирования генераций. Давно известно, что формирование пола у гаммарид детерминируется температурными условиями, при которых происходят ранние стадии онтогенеза [12]. Так, у G. duebeni при температуре ниже 5 °С продуцируются самцы, свыше 8 °С продуцируются самки, между 5 и 8 °С – и самцы, и самки.
Т.А. Бек [1] на примере популяций трех видов беломорских гаммарусов показала, что определяющее влияние на формирование пола оказывает не абсолютное значение температуры воды, а направление ее изменения: при понижающейся температуре формируются преимущественно самцы, при повышающейся – самки.
Нами была исследована сезонная динамика соотношения самцов и самок среди особей разных генераций в популяции G. oceanicus из Кольского залива. Установлено, что в каждой генерации соотношение полов неодинаково. Среди бокоплавов, вышедших из выводковых сумок в марте и мае, преобладали особи мужского пола (среднее соотношение полов в течение года составляло 3,3 и 1,5 соответственно); среди родившихся в июне – особи женского пола (среднее соотношение 0,4). Среди бокоплавов, родившихся в августе, соотношение полов в целом составляло 1:1. (табл. 1).
Таблица 1
Соотношение самцов и самок среди особей, принадлежащих разным генерациям
|
Номер генерации |
Среднее соотношение полов в генерации |
Время откладки яиц, из которых формируется генерация |
Среднемесячная температура воды на момент откладки яиц, ºС |
Время появления генерации |
Среднемесячная температура воды на момент появления генерации, ºС |
|
I |
3.3±0.5 |
декабрь |
2.4 |
март |
0.9 |
|
II |
1.5±0.1 |
февраль |
0.9 |
май |
3.6 |
|
III |
0.4±0.05 |
март |
1.7 |
июнь |
8.4 |
|
IV |
1.1±0.14 |
июнь |
8.4 |
август |
10.9 |
Была проанализирована зависимость соотношения полов в генерации от значений температур в момент выхода молоди из выводковых сумок и в момент оплодотворения яиц (рис. 3). Установлено, что существует тесная корреляционная связь между температурой воды в момент выхода молоди из марсупиумов и соотношением полов (r = 0,83): при низких значениях температуры в выводке преобладают самцы, при высоких - самки. Также проанализирована зависимость соотношения полов в генерации от направления изменения температуры воды (рис. 4). Выявлена высокая степень корреляции между направлением изменения температуры в момент оплодотворения яиц и соотношением полов (r = 0,77): когда температура воды идет на понижение, в выводке формируются преимущественно самцы; когда температура идет на повышение – самки.
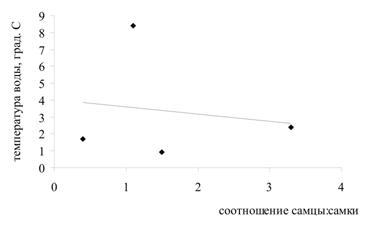
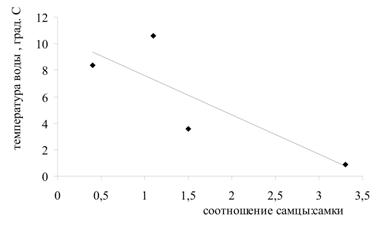
Рис. 3. Зависимость между соотношением полов в генерациях G. oceanicus и температурой воды:
а) температура воды на момент откладки яиц, из которых формируется генерация;
б) температура воды на момент выхода генерации из марсупиумов
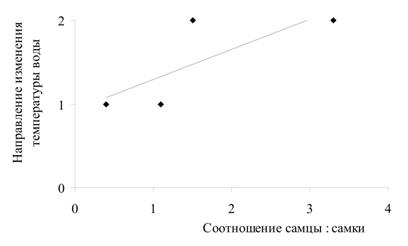
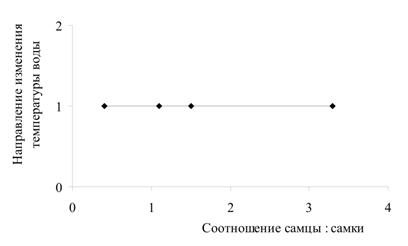
Рис. 4. Зависимость между соотношением полов в генерациях G. oceanicus и направлением изменения температуры воды:
а) направление изменения температуры воды на момент откладки яиц, из которых формируется генерация;
б) направление изменения температуры воды на момент выхода генерации из марсупиумов.
Обозначения на оси ординат: 1 – температура воды идет на повышение; 2 – температура воды идет на понижение
Некоторые исследователи в качестве ведущего фактора, влияющего на определение пола у гаммарид, называют фотопериод. В лабораторных экспериментах, проведенных Х. П. Балнеймом [9] на Gammarus dubeni, было показано, что при длинном световом дне (16 часов) в выводке формируются преимущественно самцы, при коротком (8 часов) – самки. Критической переходной точкой является длина светового дня 14 часов. Однако наши исследования показали, что в естественных условиях для Gammarus oceanicus из Кольского залива данная закономерность не подтверждается. Среди особей первой генерации, которая формируется в условиях полярной ночи и короткого светового дня, преобладают самцы, а среди особей третьей генерации, которая формируется в условиях полярного дня, преобладают самки.
Таким образом, нами показано, что формирование пола у G. oceanicus зависит от температуры воды: направление ее изменения, возможно, более важны для ранних стадий оогенеза и могут определять первичное соотношение полов, а ее абсолютные значения, по-видимому, более важны для более поздних стадий эмбриогенеза и определяют вторичное соотношение полов. По этой причине среди особей первых двух генераций, родившихся в марте и мае и пополняющих взрослую часть популяции в августе и сентябре, преобладают самцы. В октябре и ноябре начинают приобретать вторичные половые признаки особи, родившиеся в июне. Среди них преобладают самки, и соотношение полов в популяции выравнивается. В дальнейшем в популяции поддерживается приблизительно одинаковое количество самцов и самок. Незначительные отклонения от пропорции 1:1 в течение года могут возникать вследствие возрастной элиминации бокоплавов или сегрегации особей разных полов по горизонтам литорали.
Библиографическая ссылка
Икко Н.В ПОЛОВАЯ СТРУКТУРА ПОПУЛЯЦИИ GAMMARUS OCEANICUS (CRUSTACEA: AMPHIPODA) В КОЛЬСКОМ ЗАЛИВЕ // Современные проблемы науки и образования. 2010. № 1. ;URL: https://science-education.ru/ru/article/view?id=1279 (дата обращения: 27.07.2026).