



Существование гематоэнцефалического барьера существенно влияет на функции всей центральной нервной системы. Одним из морфологических субстратов этого барьера являются сосудистые сплетения [1-3]. Несмотря на довольно большое количество работ, многие стороны морфологии и физиологии сосудистых сплетений головного мозга изучены недостаточно, особенно в онтогенезе. Не прослежены эволюционные преобразования структурных компонентов сосудистых сплетений головного мозга. Отсутствует анализ морфофункционального состояния сосудистых сплетений желудочков всей циркумвентрикулярной системы [6-9]. Вместе с тем знание морфофункциональных особенностей структурных элементов гематоэнцефалического барьера может помочь раскрытию механизма защиты от проникновения фармакологических веществ, способных нарушить нормальное течение физиологических процессов в центральной нервной системе.
Целью настоящего исследования явилось изучение развития сосудистых сплетений головного мозга позвоночных животных и человека для выявления общих принципов созревания этого органа. В центре внимания было выяснение становления структуры сосудистых сплетений боковых, третьего и четвертого желудочков головного мозга в онтогенезе, начиная с ранней стадии до дефинитивного состояния, и в эволюционном аспекте.
В работе были использованы методы: анатомические (метод приготовления тотального пленочного препарата сосудистых сплетений головного мозга мышей по Сентюровой Л.Г. и Зумерову Р.А., авторское свидетельство № 1288536), гистологические: окраска гематоксилином и эозином, по Ван–Гизон, по Харту; гистохимические: окраска толуидиновым синим, по Перлсу, по Степанян Б.Г., по Шевчуку И.А. и Мельнику, по Нахласу, по Гомори и Гленнеру; по Яцковскому А.И. [10]; нейрогистологические: метод импрегнации по Куприянову В.В.; морфометрические (определяли высоту клеток хориоидного эпителия (в мкм), средний диаметр их ядер (в мкм), толщина соединительнотканной стромы (в мкм), средний диаметр отдельных звеньев микроциркуляторного русла (в мкм). Статистическую обработку полученных данных осуществляли на персональном компьютере с использованием пакета «Анализ данных» в рамках программы Microsoft Excel и BioStat 2008 Professional 5.8.4.
Результаты исследований. У круглоротых ранее считали, что в переднем мозге сосудистые сплетения отсутствуют [4; 5]. Нами обнаружено, что имеется хориоидная пластинка, к которой подрастают кровеносные сосуды. Их особенностью является наполнение форменными элементами крови. Другой особенностью структуры сплетений миноги является многослойность эпителия.
У рыб, кроме сплетения в желудочке в продолговатом мозге, встречаются сплетения и в промежуточном и в конечном мозге. Так, у осетра сосудистое сплетение переднего мозга представлено дупликатурой видоизмененной эпендимной выстилки полости желудочка. Клетки эпителия приближаются к уплощенной форме. Сосуды немногочисленны. Соединительнотканная строма практически отсутствует. Сплетения промежуточного и продолговатого мозга были представлены скоплением кровеносных сосудов с тонкой эндотелиальной выстилкой. В то же время у севрюги мы не обнаружили сосудистых сплетений в конечном мозге. А у карповых: сазана, каспийской воблы, толстолобика, лещей сосудистые сплетения развиты лишь в третьем и четвертом желудочках. Причем структура их своеобразна. Эпителиальные клетки образуют складки, которые обладают полярностью. Одна часть складки представлена клетками цилиндрической формы, другая – плоской формы. Первые имеют очень светлую, практически не окрашивающуюся цитоплазму и атипичное расположение ядра (ближе к апикальному полюсу). Такие взаимоотношения структур наблюдались у взрослых особей. Возможно, это связано со средой обитания.
Исследование онтогенеза рыб показало, что сплетения в головном мозге появляются лишь в мальковый период, а далее идет дифференцировка структурных компонентов в сторону увеличения относительных размеров.
Таким образом, по нашим данным, закладка сосудистых сплетений возникает в поздний период онтогенеза, и в дальнейшем анатомическая форма их не меняется, а лишь биометрические показатели.
Земноводные имеют сосудистые сплетения как самостоятельные образования. Структурно они оформлены складками видоизмененной эпендимы. Эпителий уже однослойный, кубический. Выражена соединительнотканная строма. В сплетениях всех желудочков наблюдается большое количество элементов крови.
Сосудистые сплетения головного мозга у рептилий хорошо выражены во всех желудочках. Особенно мощные сплетения в боковых и в четвертом желудочках. Вместе с тем общий план строения совпадает с земноводными. Однако у черепах впервые в ряду позвоночных выявляются эпиплексусные клетки.
Иную экологическую нишу занимают птицы. Закладка сосудистых сплетений головного мозга у птиц появляется на 2-й день эмбриогенеза. Сначала образуются сплетения четвертого желудочка в виде складки эпендимы и зародышевой мезенхимы. Сосудистые сплетения боковых желудочков закладываются лишь на 7-й день. Причем эпителий многорядный. Другой особенностью является наличие эпителиальных почек (зон размножения эпителия) и очагов скопления клеток в эпителиальном пласте в боковом желудочке. Мезенхима содержит многочисленные тупоначинающиеся протокапилляры. Лишь на 9-й день нами наблюдались сосудистые сплетения третьего желудочка. На третьей неделе эмбриогенеза происходит физиологическая атрофия зародышевой мезенхимы. После вылупления сосудистые сплетения продолжают морфофункциональную дифференцировку, но сохраняется заметная гетерохрония созревания разных желудочков.
У млекопитающих опережают в развитии сосудистые сплетения четвертого желудочка. Только на 15-17-й день антенатального развития появляются выпячивания в боковых желудочках медиальной стенки и сосудистой полоски третьего желудочка. Второй этап гистогенеза сосудистых сплетений начинается уже после рождения (первая – четвертая недели жизни). В этот период формируется их складчатость, но только в третьем и четвертом желудочках. Исследование пленочных препаратов показало, что формирование микрогемоциркуляторного русла продолжается и в третьем периоде (начиная с 5-й недели до дефинитивного состояния).
Наивысшее развитие сосудистые сплетения достигают у человека. Сплетения продолговатого и промежуточного мозга образуются путем инвагинации однослойной крыши. В то время как сплетения боковых желудочков (конечного мозга) развиваются из медиальной части из многоклеточного матрикса. Раньше всего появляется закладка сосудистого сплетения четвертого желудочка. В 7 недель сплетения боковых желудочков представлены короткими первичными ворсинами. К 8-й неделе появляются вторичные выпячивания. В этот период на некоторых участках эпителий имеет псевдомногослойный характер. Строма представлена мезенхимой, содержащей гемоцитобласты и ангиобласты. Уже к 15 неделям сосудистые сплетения занимают большую часть желудочков. Ядра эпителиоцитов располагаются ближе к апикальному полюсу. Дальнейшие морфофункциональные изменения после 16 недель касаются не только перемещения ядра эпителиоцита к базальному полюсу, но и формирования в сосудистых сплетениях бокового и четвертого желудочков особого участка - гломуса.
Обсуждение результатов. Формирование структур гематоэнцефалического барьера играет существенную роль для обеспечения нормального функционирования головного мозга [3-5]. Сравнительное исследование сосудистых сплетений головного мозга у круглоротых, рыб, амфибий, рептилий, птиц, млекопитающих и человека позволило выявить закономерности их морфогенеза. Процессы дифференцировки наблюдаются во всех структурных компонентах: эпителии, соединительнотканной строме, микроциркуляторном русле.
Основными закономерностями филогенетических изменений строения сосудистых сплетений головного мозга позвоночных животных являются: прогрессивное усложнение их организации, приобретение ими однотипного строения с многоярусным ветвлением ворсин во всех желудочках головного мозга у человека.
Анализ морфометрических данных на примере млекопитающих (рис. 1). На гистограмме видно, что эпителиоциты хориодных сплетений (хориоэпителиоциты) четвертого желудочка во все сроки развития по размерам больше, чем аналогичная структура в боковых и третьем желудочке. Очевидно, это обусловлено большей функциональной нагрузкой у млекопитающих этого отдела циркумвентрикулярной системы головного мозга. Сосудистые сплетения боковых и третьего желудочков статистически достоверных различий по морфометрическим показателям не обнаружили.
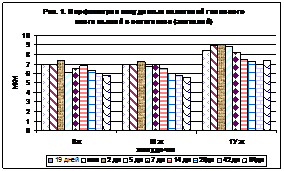
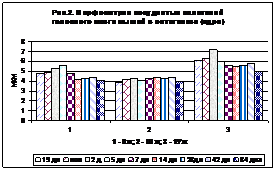
Рис. 1. Рис. 2.
На рис. 2 у млекопитающих также заметно больше объем ядер хориоэпителиоцитов четвертого желудочка.
В то же время ядра всех эпителиоцитов сосудистых сплетений можно поделить на: функционально активные (при окраске по Яцковскому А.И. - синие), промежуточные (сине-красные) и функционально неактивные (красные) красные. Так, на рис. 3 видно, что во всех желудочках идет увеличение количества активных ядер к 20 дням постнатальной жизни. Причем в большей степени в боковом желудочке. Также отчетливо видно, что после 42 дней постнатальной жизни активность ядер резко падает.
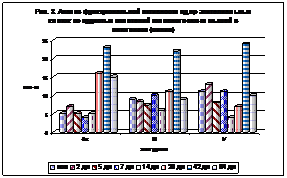
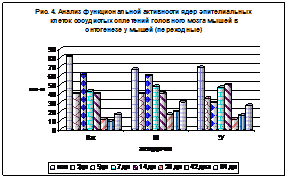
Рис. 3. Рис. 4.
Что касается количества переходных ядер (рис. 4), то видно, что больше наблюдается в молодом возрасте.
Обращает на себя внимание (рис. 5), что большая часть ядер эпителиоцитов сосудистых сплетений всех желудочков головного мозга функционально неактивна.
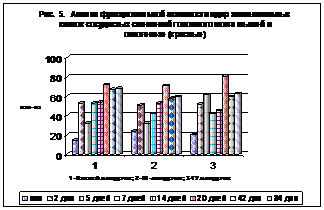
Рис. 5.
Заключение. Исследование сосудистых сплетений головного мозга позвоночных в сравнительном аспекте позволило установить закономерности, свидетельствующие об их морфофункциональных особенностях в онто- и филогенезе позвоночных. Процессы развития можно проследить во всех основных морфологических структурах, в частности в хориоэпителиоцитах.
Рецензенты:Молдавская А.А., д.м.н., профессор кафедры анатомии человека Астраханской государственной медицинской академии, г. Астрахань.
Фельдман Б.В, д.б.н., доцент, зав. кафедрой ботаники, фармакогнозии и фармацевтической технологии Астраханской государственной медицинской академии, г. Астрахань.
Библиографическая ссылка
Сентюрова Л.Г. МОРФОФУНКЦИОНАЛЬНОЕ СТАНОВЛЕНИЕ СОСУДИСТЫХ СПЛЕТЕНИЙ ГОЛОВНОГО МОЗГА ПОЗВОНОЧНЫХ ЖИВОТНЫХ И ЧЕЛОВЕКА // Современные проблемы науки и образования. 2014. № 2. ;URL: https://science-education.ru/ru/article/view?id=12469 (дата обращения: 29.06.2026).