



Введение
Целью воспроизводства лесов является рациональное использование лесных земель, оптимизация формационной и возрастной структуры лесов, повышение их продуктивности, устойчивости и качества, сохранение и восстановление растительного биоразнообразия, улучшение экологической обстановки [1]. Достижение данной цели возможно при применении в лесах различной категории целевого назначения определённых способов лесовосстановления. Поэтому задача современного лесоводства заключается в выборе способа лесовосстановления для конкретных условий [3,5,6].
Цель данного исследования – изучение лесных культур сосны обыкновенной в биогруппах и бороздах, сравнение их хода роста и оценка с экологической точки зрения. Материалы исследования были собраны в полевые сезоны 2011–2013 годов на территории Тамбовской области. Леса региона образуют три крупных лесных массива: Цнинский – 252,6 тыс. га (67 %), Иловайский – 51 тыс. га (14 %) и Воронинский – 48,2 тыс. га (13,0 %). Остальная площадь лесов рассредоточена в южной и юго-западной части области среди сельскохозяйственных земель и представлена отдельными урочищами и колками [5].
Для типа условий местопроизрастания С3 на территории Цнинского лесничества характерно вытеснение главной породы (сосны обыкновенной) осиной [1,2,3]. Поэтому в 60–70 гг. 20 века здесь была произведена посадка культур сосны обыкновенной биогруппами. На 1 гектар при таком способе посадки приходилось 250–280 биогрупп. Примерное количество 2-х летних сеянцев сосны обыкновенной на одну древесную группу – 25–28.
В процессе исследований на территории Цнинского лесничества были произведены измерения основных биометрических параметров биогрупп и посадок в борозды сосны обыкновенной разного возраста (табл.1,2).
Таблица 1. Деревья центрального ряда биогрупп
|
№ ПП |
Dср±m |
δ |
Нср±m |
δ |
Количество деревьев |
ТУМ |
Возраст, лет |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
1 |
2,5±0,17 |
1,15 |
1,6±0,1 |
0,58 |
42 |
С3 |
6 |
|
2 |
2,3±0,2 |
0,9 |
1,6±0,1 |
0,6 |
31 |
С3 |
6 |
|
3 |
7,6±0,62 |
3,6 |
8,2±0,45 |
2,6 |
30 |
С3 |
15 |
|
4 |
9,6±0,42 |
2,4 |
9,5±0,37 |
2,1 |
32 |
С3 |
21 |
|
5 |
12,3±0,7 |
4 |
10,9±0,53 |
2,9 |
30 |
С3 |
21 |
|
6 |
13,8±0,89 |
4,9 |
12,3±0,57 |
3,1 |
30 |
С3 |
24 |
|
7 |
20±0,6 |
3,4 |
20±0,28 |
1,6 |
32 |
С3 |
39 |
|
8 |
21,5±0,58 |
3,3 |
21,1±0,28 |
1,6 |
32 |
С3 |
41 |
|
9 |
24,7±1,1 |
6,0 |
23±0,65 |
3,7 |
32 |
С3 |
46 |
Деревья сосны обыкновенной центральных рядов биогрупп более стабильны по высоте, чем по диаметру. На пробных площадях 1–9 (в пределах возрастных групп от 6 до 49 лет) значения стандартных отклонений по высотам меньшие. Максимальное значение δ принимает в возрасте 46 лет, минимальное – в 6 лет (табл.1). Изменение данной статистической величины по всем пробным площадям не является линейным.
Таблица 2. Деревья крайних рядов рассматриваемых биогрупп
|
№ ПП |
Dср±m |
δ |
Нср±m |
δ |
Количество деревьев |
ТУМ |
Возраст, лет |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
1 |
1,9±0,07 |
0,8 |
1,4±0,06 |
0,5 |
42 |
С3 |
6 |
|
2 |
2,2±0,1 |
0,9 |
2,3±0,1 |
0,8 |
31 |
С3 |
6 |
|
3 |
8,8±0,7 |
4,1 |
8,3±0,52 |
3,1 |
30 |
С3 |
15 |
|
4 |
11,1±0,7 |
4 |
10,6±0,6 |
3,4 |
32 |
С3 |
21 |
|
5 |
10,7±0,65 |
4,2 |
10,1±0,51 |
3,3 |
30 |
С3 |
21 |
|
6 |
11,3±0,47 |
2,8 |
11,1±0,32 |
1,9 |
30 |
С3 |
24 |
|
7 |
18,3±0,75 |
4,6 |
19,0±0,76 |
4,7 |
32 |
С3 |
39 |
|
8 |
20,6±0,62 |
3,5 |
22,8±0,35 |
2,0 |
32 |
С3 |
41 |
|
9 |
26,7±0,98 |
6,1 |
24,5±0,64 |
4,0 |
32 |
С3 |
46 |
Для деревьев крайних рядов биогрупп характерно: диаметр максимально варьирует на ПП9 (возраст 46 лет); максимальное отклонение высоты отмечено в 39-летнем возрасте (δ принимает значение равное 4,7). Наиболее минимальные значения стандартное отклонение принимает на участках ПП1, ПП2 (шестилетние молодняки) (табл. 2). Здесь пределы изменения величины стандартного отклонения деревьев биогрупп по диаметрам и высотам близки.
Распределение деревьев в биогруппах по диаметрам и высотам изменяется с возрастом. Минимальное значение статистическая величина стандартного отклонения принимает в обоих случаях в молодняках. Однако и для деревьев крайних рядов не характерны изменения δ как линейной величины. Структура рассматриваемой ценопопуляции сосны обыкновенной в пределах определённой возрастной группы формируется под воздействием биотических факторов, особую роль среди которых играют конкурентные взаимоотношения между особями одного вида.
Показателями остроты конкурентных отношений могут служить данные по распределениям деревьев биогрупп по классам роста Крафта. Основной полог лесных фитоценозов, созданных по бороздам, образуют деревья II класса роста (до 40 % по числу и до 70 % по запасу древесины); их кроны имеют нормальное развитие. Деревья I класса роста несколько выше средних показателей, их кроны более низко опущенные и занимают до одной трети протяженности всего ствола; количество таких деревьев около 5 %. Деревья III класса несколько ниже среднего показателя, кроны их более узкие, чем у деревьев II класса, но полностью входят в основной — верхний
полог древостоя. Деревья IV класса имеют очень узкую (IVa) или однобокую (IV6) крону и по высоте ниже деревьев III класса [4].
В зависимости от состояния деревьев биогрупп и с учётом их биометрических параметров было произведено разделение деревьев отдельно для центральных и крайних рядов на классы роста Крафта (табл.3,4).
Таблица 3. Распределение деревьев центральных рядов по классам роста Крафта
|
№ |
Возраст, лет |
Размещение в площадках |
К-во деревьев |
Распределение по классам Крафта, % |
|||
|
Кл1 |
Кл2 |
Кл3 |
Кл4 |
||||
|
1 |
6 |
3,5*2 |
42 |
36 |
24 |
31 |
9 |
|
2 |
6 |
3*2 |
31 |
19 |
36 |
23 |
23 |
|
3 |
15 |
3,8*2 |
30 |
37 |
13 |
23 |
27 |
|
4 |
21 |
2,4*2 |
32 |
67 |
3 |
18 |
12 |
|
5 |
21 |
3*2 |
30 |
60 |
- |
33 |
7 |
|
6 |
24 |
3*2 |
30 |
63 |
6 |
3 |
28 |
|
7 |
39 |
3*2 |
34 |
94 |
- |
6 |
- |
|
8 |
41 |
2*2 |
32 |
91 |
6 |
3 |
- |
|
9 |
46 |
2*2 |
32 |
73 |
- |
12 |
15 |
Деревья 4 классов роста Крафта отмечены на ПП1, ПП2,ПП3, ПП4, ПП6 и ПП9 (возраст 6,15,21,24 и 46 лет соответственно). На остальных участках отсутствуют деревья 2 или 4 классов роста. Так, например, в возрасте 39 лет (пробная площадь № 7) характерно наличие деревьев 1 и 3 классов роста Крафта. Для биогрупп с деревьями 1-го класса возраста характерно преобладание деревьев 1 и 2-го классов роста Крафта. К более старшему возрасту увеличивается доля деревьев первого класса роста Крафта (более 50 %) (табл. 3). Дифференциация деревьев по классам роста Крафта зависит также и от размера биогрупп. В наиболее крупных биогруппах встречаются деревья всех четырех классов роста по Крафту (причем доля деревьев 3-го и 4-го классов значительна и составляет в сумме не менее 50 %).
Таблица 4. Распределение деревьев крайних рядов по классам Крафта
|
№ |
Возраст, лет |
Размещение в площадках |
К-во деревьев |
Распределение по классам Крафта, % |
|||
|
Кл1 |
Кл2 |
Кл3 |
Кл4 |
||||
|
1 |
6 |
3,5*2 |
68 |
36 |
10 |
31 |
23 |
|
2 |
6 |
3*2 |
53 |
13 |
42 |
15 |
30 |
|
3 |
15 |
3,8*2 |
36 |
44 |
3 |
31 |
22 |
|
4 |
21 |
2,4*2 |
32 |
50 |
- |
25 |
25 |
|
5 |
21 |
3*2 |
31 |
39 |
- |
32 |
29 |
|
6 |
24 |
3*2 |
35 |
80 |
- |
11 |
9 |
|
7 |
39 |
3*2 |
38 |
76 |
- |
13 |
11 |
|
8 |
41 |
2*2 |
32 |
72 |
- |
25 |
- |
|
9 |
46 |
2*2 |
38 |
52 |
- |
24 |
24 |
Крайние ряды характеризуются другим распределением по классам роста Крафта: деревья 1–4 классов роста представлены на ПП1-ПП3 (возраст 6–15 лет). В возрастном интервале от 21 года до 46 лет отсутствуют деревьев второго класса роста Крафта. Доля деревьев 3 и 4 классов роста Крафта значительна (в сумме от 20 до 61 %). Этот факт может быть объяснен не только размерами биогрупп и внутривидовыми конкурентными взаимодействиями, но и межвидовой конкуренцией (табл. 4).
Для центральных рядов в более старшем возрасте характерно наличие значительного количества деревьев 2-го класса роста Крафта. Деревья, произрастающие в крайних рядах, характеризует более сложная дифференциация деревьев по классам роста, и значительная доля их отнесена к 4 классу роста. Такое распределение деревьев по классам роста Крафта обеспечивается функционированием естественных механизмов, направленных на поддержание потенциала сообщества.
Таким образом, деревья в биогруппах имеют структуру насаждения, отличную от деревьев по бороздам. Причиной таких расхождений является внутривидовая конкуренция, которая определяется положением видов в сообществе, то есть их экологическими нишами. Конкуренция происходит в тех случаях, когда взаимодействие между двумя или несколькими особями или популяциями оказывает неблагоприятное влияние на рост, выживание и приспособленность каждой особи. В типичном случае это происходит из-за недостатка ресурса. То есть на распределение деревьев по классам роста Крафта влияет первичная площадь биогрупп или площадь питания на 1 сеянец сосны обыкновенной. Если потребность в ресурсе уравнивается, то особи конкурируют меньше. Внутривидовая конкуренция способствует, с одной стороны, расширению ниши, а с другой – увеличению разнообразия. Если в том или ином сообществе есть свободные экологические ниши, то существующие здесь виды получают некие дополнительные ресурсы (Джиллер, 1988).
На основании экспериментальных и справочных данных построены графики хода роста деревьев в биогруппах по диаметру и высоте в сравнении с бороздными культурами (рисунки 1 и 2).
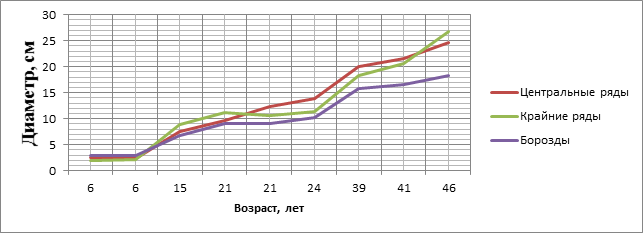
Рисунок 1. Ход роста по диаметру деревьев, созданных биогруппами и по бороздам
В возрасте 6 лет больший диаметр имеют деревья, произрастающие по бороздам. После смыкания деревьев в биогруппе (7–9 лет) характеризуются большим приростом по диаметру. Максимальное отклонение от посадок в бороздах биогруппы имеют в возрасте 46 лет, минимальное – в возрасте 24 лет, а для деревьев центральных и крайних рядов – в возрасте 21 года (рис.1).
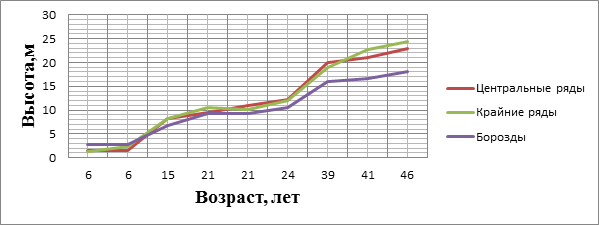
Рисунок 2. Ход роста по высоте деревьев биогрупп в сравнение с эталонным насаждением
В возрасте 6 лет деревья сосны обыкновенной по бороздам обгоняют по высоте посадки в биогруппах. Но начиная с 10–15 лет прослеживается постоянное нарастающее преобладание роста по высоте в биогруппах. При этом рост в высоту более выражен в центральных рядах в период интенсивного самоизреживания куртин (биогрупп) (20–40 лет). Ко времени смыкания биогрупп положение деревьев в них не так сказывается на их энергии роста. Контакт с древесными растениями фона увеличивается, начинается становление сложного по составу древостоя, соответствующего типу лесорастительных условий.
Выводы:
1. Способ создания лесных культур (способ лесовосстановления) оказывает большое влияние на формирование лесного фитоценоза в будущем.
2. Особую роль играют конкурентные механизмы, в том числе внутривидовые.
3. Лесные культуры в биогруппах и в бороздах отличаются по биометрическим параметрам и структуре насаждения. Деревья в биогруппах характеризуются большим приростом по высоте и диаметру с момента смыкания крон.
4. Устойчивые экологические функции выполняют насаждения с более высокими показателями прироста по высоте и диаметру. Следовательно, биогруппы выполняют свои экологические функции более стабильно и эффективно.
Рецензенты:
Арефьев Ю.Ф., д.б.н., профессор, профессор кафедры экологии, защиты леса и лесного охотоведения Федерального государственного бюджетного образовательного учреждения высшего профессионального образования «Воронежской государственной лесотехнической академии», г. Воронеж.
Харченко Н.А., д.б.н., профессор, профессор кафедры экологии, защиты леса и лесного охотоведения Федерального государственного бюджетного образовательного учреждения высшего профессионального образования «Воронежской государственной лесотехнической академии», г. Воронеж.
Библиографическая ссылка
Семёнов М.А. ОСОБЕННОСТИ ФОРМИРОВАНИЯ СТРУКТУРЫ ИСКУССТВЕННЫХ ФИТОЦЕНОЗОВ ПРИ СОЗДАНИИ ЛЕСНЫХ КУЛЬТУР БИОГРУППАМИ // Современные проблемы науки и образования. 2013. № 6. ;URL: https://science-education.ru/ru/article/view?id=11181 (дата обращения: 17.07.2026).