



ВВЕДЕНИЕ
При решении проблемы безопасности космических полетов и, в частности, полета к Марсу необходимы более глубокие представления о механизмах воздействия различных физических факторов полета на биообъекты и, в частности, человека. Только знания этих механизмов позволит прогнозировать возможные новые эффекты опасности и предлагать меры защиты от них. Особенно актуально задача выяснения механизма действия адронов высоких энергий, γ-радиации и слабых, низкоинтенсивных воздействий физических и химических факторов, на что обращали внимание ранее многие исследователи [1,9,10,12].
Проведённый нами анализ биологических экспериментов в космосе [1,3-5,11] и собственных результатов показал, что зарегистрированные низкие дозы космической радиации вызывали статистически достоверный биологический эффект. Эти результаты пытались объяснить действием невесомости и радиацией. Однако модельные эксперименты, проведенные в земных условиях и в космическом пространстве, не подтвердили этого предположения. При применении микродозиметрического анализа для изучения наблюдаемых биологических эффектов в космосе и на ускорителях удовлетворительного объяснения также не было получено.
Предложенные нами ранее магнитно-резонансный механизм (МРМ) [9,10] слабых воздействий и концепция фундаментальной роли реликтового излучения (РИ) Вселенной применена к анализу и прогнозированию новых эффектов в межпланетных полетах и к обеспечению безопасности космонавтов.
МАТЕРИАЛЫ И МЕТОДЫ
В нашем эксперименте семена Lycopersicon esculentum Mill. экспонировались на космическом орбитальном комплексе «МИР» в течение шести лет. После этого полётные семена выдерживались некоторое время в лабораторных условиях (для элиминации обычных одиночных, точечных повреждений). Затем с семенами были проведены генетические и анатомо-морфологические исследования.
Опытные (полётные) семена замачивались на 2 часа в водопроводной воде. После этого семена выдерживали в 1% растворе марганца в продолжение 20 минут, затем в течение двух минут отмывали в водопроводной воде. Затем семена подсушивались при комнатной температуре в течение двух часов. После этих обработок семена высевались в приготовленную садовую землю (1/2 торфа, 1/2 дерновой земли и 100 грамм золы на 8 кг почвы). Приготовленная почва перед посадкой семян прокаливалась в газовой печи 20 минут. В течение 70 суток семена анализировались в лабораторных условиях. Высев в полевые условия производился в первых числах мая в приготовленные грядки под плёнку. В процессе прорастания и развития семян в лабораторных условиях (70 суток) анализировались следующие показатели: выживаемость, «оживление», летальность, физиологические показатели растений, эмбриональные уродства в эмбриогенезе и мутагенез. В открытом грунте исследовался летальный эффект, морфогенез, созревание и анализ плодов. Обработка семян ВБИ производилась по методу [10], который позволяет оживить условно-летальные семена томатов и тем самым выявить те летальные мутации, которые находились в скрытом «летальном» состоянии.
В чашках Петри выращивалась культура дрожжей Sporobolomyces alborubescens Derx. с толщиной монослоя 2 мм. Затем эти чашки облучались дозами γ-излучения 137Cs 1.10-2 – 5.10-8 сГр. На облученные чашки помещали сухие семена на 6 часов. В течение этого времени семена (90 шт.) подвергались воздействию ВБИ. В результате экспериментов было определено, что оптимальная доза ВБИ составляла 1,2.10-4 сГр. Природный радиационный фон составляет 1.10-6 сГр за 6 часов. Статистическую обработку экспериментальных результатов проводили по непараметрическим критериям. Достоверность различий между двумя выборками совокупностей определяли по серийному и Колмогорова-Смирнова критериям [7]. Различия считали статистически значимыми при Р<0,05 и Р<0,01.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Прорастание полетных семян томатов (без ВБИ) происходило на 14 сутки проращивания, в то время как в контрольных семенах без воздействия факторов космического полёта - на 5-ые сутки. Обработанные ВБИ полётные семена начинали проклёвываться на 11 сутки.
Всхожесть семян в полётном варианте (без воздействия ВБИ) составляла 26,8%, а в контрольном варианте 58,3%, следовательно, всхожесть в опыте была в 2 раза ниже, чем в контроле.
Плоды растений, выросших в F1 поколении из полетных семян, были тёмно-красного цвета, вес плодов колебался от 30 до 100 г и более, мякоть плода буро-темно-красного цвета, плоды были 2-х, 3-х, 4-х и 7-ми камерные.
В опытных плодах в F1 поколении образование семян проходило с большими нарушениями. Обобщенные результаты по опытным плодам показали: нормальных семян было 36,2% - 52,8% (в разных повторностях), а абортированных (не сформировавшихся) 47,2% - 63,8%.
В контрольных семенах, выдержанных 6 лет и более без воздействия ФФКП и без воздействия ВБИ, полностью отсутствовали абортированные семена. Наличие абортированных семян в полётных вариантах плодов F1 поколения указывает на несбалансированность генетического аппарата зародышей семян, которая вызвана крупными перестройками генетического материала, - делеции (крупные и протяженные), вставки. Аналогичные генетические изменения были обнаружены в исследованиях на бактериофаге T4Br+ [1,8,12].
Нами был проведен высев в два приёма 41-ой штуки опытных семян томатов (21 шт. и 20 шт.) с разницей 18 суток без обработки ВБИ. Взошло всего 11 семян, что составляло общую всхожесть 26,8%. .
Была поставлена задача: выявить скрытые изменения биолого-генетического характера, применяя разработанный нами ранее метод оживления условно-летальных семян (клеток), так называемый оживляющий феномен ВБИ [6,10]. На основе этого метода мы попытались выявить специфические повреждения у семян, индуцированные действием ФФКП.
Максимум всхожести, обусловленной оживляющим действием ВБИ, наблюдался при 1,4.10-4 сГр, он составлял 75% прорастания в опыте и 26,8% - без воздействия ВБИ.
Следовательно, истинный эффект оживляющего действия ВБИ равнялся 48,2% и, таким образом, выяснилось, что практически 50% всех полётных семян имели скрытые «летальные» генетические повреждения, индуцированные действием ФФКП. Оставшиеся 25% полетных семян вообще полностью потеряли способность к оживлению и были абсолютно-летальными. Таким образом, анализ экспериментальных результатов показал, что 48,2% «оживленных» семян имели скрытые повреждения и 25% семян были абсолютно-летальными, т.е. отчетливо был выявлен оживляющий феномен ВБИ.
Далее нами была проанализирована специфичность мутагенного действия ФФКП на семена томатов
Использование нами оживляющего феномена ВБИ увеличило в 3 раза выявление скрытых мутаций, которые были первично индуцированы действием ФФКП. Растения с такими морфологическими мутациями названы были нами впервые как бессемядольные, безлиственные «пеньки», а более точно - гипокотельные пеньки (ГКП) (рис. 1, 2 — фотокомпьютерная обработка). Такие мутанты, полученные из условно-летальных семян посредством оживляющего феномена ВБИ, никогда не наблюдались как в опытных, так и в лабораторных контрольных исследованиях в земных условиях. ГКП-мутанты и их продвинутые мутации не давали полноценных растений. Они, как правило, погибали после 1,5-2 месяцев роста в лабораторных условиях. ГКП-мутанты обычно достигали высоты 7 - 25 мм, при диаметре стебля «пенька» до 1,2 мм. Мутанты ГКП – ярко зелёные, слабо опушенные, длина волоска 0,1-0,2 мм, снизу 5 мм от почвы – неопушенные, верх «пенька» подобен струпу, имеются болячки бело-серого цвета. У некоторых ГКП-мутантов и продвинутых ГКП-мутантов вверху наблюдались ороговевшие и засыхающие 2-3 площади диаметром 0,2 мм (засохшие точки роста семядолей и первичных листьевых стеблей).
Таких ГКП мутантов и продвинутых ГКП мутантов мы получили 16 штук. Необходимо отметить, что при составлении генетических карт томатов, начатом ещё в 1933 г. отдельными исследователями, а также Международной ассоциацией по генетике томатов (Tomato Genetics Cooperative – TGC) с 1950 г. по настоящее время было исследовано более 2000 мутаций, включая и спонтанные [2], но мутаций, подобных полученному нами ГКП мутанту, обнаружено не было. При этом индукция мутаций проводились всеми возможными физико-химическими мутагенами широкого диапазона спектра действий. Следовательно, впервые нами получены специфические ГКП подобные мутанты, индуцированные ФФКП.
| | |
| Рис. 1. Фото – (компьютерная обработка). Гипокотельные пеньки-мутанты (ГКП) у растений томатов, индуцированные действием ФФКП при шестилетнем полёте на космическом орбитальном комплексе «МИР» и выявленные посредством «оживляющего» феномена ВБИ. | Рис. 2. Фото – (компьютерная обработка). Семядольные мутанты (продвинутая ГКП мутация) с семядольным ростом (СР) у растений томатов, индуцированные действием ФФКП на космическом орбитальном комплексе «МИР» с 6-летней экспозицией и выявленные посредством «оживляющего» действия ВБИ. |

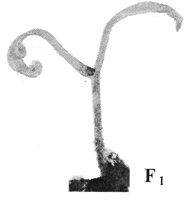
Можно предположить, что у ГКП-мутантов программа развития в процессе онтогенеза у растения заблокирована на стадии выброса семядолей и листьев, возможно, что эта мутация расположена во второй хромосоме [2] (по аналогичным мутантам). Остальные мутации семян следующие: булава, очень похожая на ГКП мутацию, но только с двумя семядолями; ГКП мутация, но только с семядольным ростом; ГКП с двумя долями и только с одним листом (табл. и фото 1,2), скорее всего, это, по-видимому, дальнейшая онтогенетическая модификация истинного ГКП-мутанта. Подобные мутации у семян растений, продвинутых в начальной стадии онтогенеза, - ещё дополнительное доказательство в пользу специфичности мутагенного воздействия ФФКП.
Таблица 1. Распределение оживленных, посредством вторичного биогенного излучения, морфологических мутаций у проростков опытных семян Lycopersicon esculentum Mill., подвергнутых шестилетнему воздействию физических факторов космического пространства.
| Дозы ВБИ в сГр | Количество морфологических мутаций проростков семян (в штуках) | ||||||||
| ГКП | БВ | БуВ | СР | 2СЛ | СЖ | 2СЛМ | СкЛ | ||
| 1,2.10-1 | ВБИ | 2 |
|
|
|
|
|
|
|
| 1,2.10-2 | ВБИ | 1 |
|
|
|
|
|
|
|
| 1,2.10-3 | ВБИ | 3 |
|
| 1 | 1 |
|
|
|
| 1,2.10-4 | ВБИ | 1 |
| 2 |
|
|
| 1 |
|
| 1,4.10-4 | ВБИ | 1 |
|
|
|
| 2 | 1 |
|
| 1.10-6 | ВБИ |
|
|
|
|
|
| 2 |
|
| 5.10-8 | ВБИ |
|
|
|
|
|
|
|
|
| Коп 1.10-6 | 1 |
|
|
|
|
| 1 | 1 | |
| Клаб. 1.10-6 |
| 1 |
|
|
|
|
| 1 | |
Обозначения мутаций: ГКП – гипокотельные «пеньки»; БВ – бокаловидные; БуВ – булавовидные; ГКП-СР – семядольный рост; ГКП-2СЛ – две семядоли и один лист; СЖ – семядоли желтые; ГКП-2СЛМ – две семядоли и маленький лист; СкЛ – скрученные листья.
На основе разработанного нами магнитно-резонансного механизма [9,10] и поглощения реликтовых компонент (переносчики всех известных четырех физических фундаментальных взаимодействий) частицами, атомами, молекулами объясняется возникновение первичных и вторичных поляризованных излучений. Это происходит при компенсации квантовых орбит электронов в атомах за счет реликта посредством ядерно-магнитного резонанса (ЯМР) и электроно-парамагнитного резонанса (ЭПР), когда возникает первичное поляризованное излучение (ППИ). В биологических объектах естественно присутствует in vivo все необходимые физические вышеперечисленные параметры, которые требуются для осуществления названных взаимодействий, при определенном накоплении ППИ они переходят в новое качество – ВБИ, при котором появляются вторичные поляризации электронов. Сформировавшееся ВБИ передает затем сигнал через водную межклеточную среду (свободную воду) рецепторам, потом через клеточный мембранный потенциал (МП) и далее мигрирует по конденсированным структурам ДНК, РНК, белков в клетках и тканях.
Реликтовое излучение (РИ) постоянно и везде в материальном мире участвует в поддержке устойчивого равновесного состояния природных систем: от стационарных орбит электрона в атоме до стационарного мембранного клеточного потенциала в организме. Энергия, теряемая электроном как заряженной частицей, движущейся с ускорением, компенсируется энергией резонансно поглощаемых реликтовых фотонов. Устойчивость МП, являющегося критериальным, интегральным показателем нормы и патологии клетки [5,9], при его, казалось бы неизбежном понижении за счет пассивного диффузионного транспорта ионов, поддерживается путем резонансного поглощения энергии РИ. Накачка энергии может происходить и при различных химических реакциях по механизму химической поляризации ядер и электронов, ионов и свободных радикалов. Очень важные для клетки химические процессы, происходящие в мембране, связаны с образованием энергии и изменением липидного состава мембран. Свободно-радикальные процессы перекисного окисления липидов зависят от фазового (физического) состояния мембран и ФФКП с последующей цепочкой воздействий на нуклеино-белковый комплекс клетки. Обнаруженная роль РИ носит фундаментальный, универсальный характер, поскольку поглощение соответствующих компонент реликта обеспечивает возможность всех типов взаимодействий, как, например, поглощение скоррелированных пар фотонов (типа пары фотонов в парадоксе Эйнштейна-Подольского-Розена) – электромагнитных взаимодействий, пары нейтрино-антинейтрино (концентрация их составляет около 200 пар в cм3 и средней энергией 10-4 эВ) – слабых взаимодействий (ответственных за β-распад), не считая адронных и гравитационных взаимодействий. Первая составляющая РИ – природная, анизотропная, поляризованная – отвечает за изменчивость вида, т.е. за мутагенез (который как мы показали, имеет пороговую дозовую зависимость). И все биологические объекты излучают эту поляризационную составляющую, по частоте совпадающую с РИ (как отмечалось выше), это и есть ВБИ, которое мигрирует и имеет коэффициент усиления 104. Именно этот феномен участвует в оживлении условно-летальных клеток при их соседстве с нормальными здоровыми клетками, содержащими поляризованную составляющую (интересно, что оживление имеет беспороговую дозовую зависимость). Вторая составляющая РИ – постоянная, изотропная, неполяризованная – обеспечивает устойчивость вида, т.е. его консерватизм. Просматривается аналогия между концепцией реликтового излучения с ее двумя фундаментальными составляющими и эволюционной теорией, в которой такие понятия, как «изменчивость видов» (мутация) и «устойчивость видов» («оживление») являются основой учения.
ЗАКЛЮЧЕНИЕ
Исследования «оживления» семян посредством ВБИ и последующее изучение генетических и физиологических изменений у растений, выросших из таких семян, которые перед этим были длительное время экспонированы в условиях космического полёта, обнаружили ряд закономерностей.
Максимум оживляющего действия ВБИ на опытных семенах томатов составлял 48,2%, то есть практически 50% всех исследуемых семян, подвергнутых влиянию ФФКП, имели скрытые «летальные» повреждения.
Абсолютно-летальных семян в опытном варианте было 25%. Количество всех скрытых повреждений в семенах достигало 75%.
Максимум выхода генетических мутаций был в 3 раза выше у семян, подвергнутых воздействию ФФКП с последующей обработкой ВБИ.
Последующая гибель «оживленных» растений с условно-летальными мутациями, индуцированными действием ФФКП, в лабораторных условиях составляла не более 15% в течение 2,5 месяцев роста.
Впервые обнаружен специфический гипокотельный пеньковый тип мутаций (ГКП) с модификациями, которые первично были индуцированы действием ФФКП.
Впервые разработана концепция реликтового излучения, которая ответственна за образование ВБИ, за изменчивость (мутагенез) и за устойчивость (консерватизм) биологического вида.
СПИСОК ЛИТЕРАТУРЫ:
1. Акоев И.Г., Юров С.С // Биофизические основы космической радиации и излучения ускорителей. Проблемы космической биологии. Т. 60. Ленинград. «Наука». 1989. С. 7.
2. Жученко А.А., Балашова Н.Н., Король А.Б., Самовол А.П., Грати В.Г., Кравченко А.Н., Кравченко А.Н., Добрянский В.А., Смирнов В.А., Бочарникова Н.И. // Эколого-генетические основы селекции томатов. Отв. ред. Лысиков В.Н. Кишинёв. «ШТИИНЦА» 1988. С. 145.
3. Иванов В.И., Дмитриевский И.М. // Проблемы космической биологии. Т. 60. Ленинград. «Наука». 1989. С. 80.
4. Ковалёв Е.Е., Брилль О.Д. // Вопросы биологического действия и дозиметрии тяжёлых заряженных частиц и адронов высоких энергий. Пущино. Изд-во АН СССР. 1984. С. 144.
5. Кожокару А.Ф., Кожокару Н.Л., Бурковецкая Ж.И. // Биомедицинские технологии и радиоэлектроника. 2006. № 8-9. С. 58.
6. Кузин А.М., Юров С.С., Ревин А.Ф. // Радиационная биология. Радиоэкология. 1997. Т. 3. вып. 5. С. 275.
7. Урбах В.Ю. // Математическая статистика для биологов и медиков. Москва. Изд-во АН СССР. 1963. С.279.
8. Юров С.С., Ливанова И.А., Смирнова Е.Н., Стаканов В.А., Маренный А.М., Попов В.И., Нечитайло Г.С. // Успехи космической биофизики. Пущино. Изд-во АН СССР. 1978. С.38.
9. Дмитриевский И.М. // Полигнозис. №2. 2000. С.38.
10. Dmitrievsky I.M., Yurov S.S., Leontyeva G.A. // In Genetic Consequences of emergency radiation situations: International conference. Moscow, Russia, 10 – 13 June, 2002. Proceedings – Moscow. Publishing House of Russian Peoples Friendship. 2002. P. 59.
11. Nechitailo G.S., Мashinsky A.L. // Space Biology, Studies at the orbital stations, Mir Publishers. Moscow. 1993. Р.603.
12. Yurov S.S., Akoev I.G., Akhmadieva A.Kh., Livanova I.A., Leontyeva G.A., Marennyi A.M., Popov V.I. // Life science and space research. Vol. XVII. Oxford and New York. Ed. by R. Holmquist, Pergamon Press. 1979. P. 129.
Библиографическая ссылка
Юров С.С, Кожокару А.Ф, Дмитриевский И.М, Нечитайло Г.С ГЕНЕТИКО-ФИЗИОЛОГИЧЕСКИЕ И ФИЗИКО-ХИМИЧЕСКИЕ ИССЛЕДОВАНИЯ LYCOPERSICON ESCULENTUM MILL. , ВЫРАЩЕННЫХ ИЗ СЕМЯН, ЭКСПОНИРОВАННЫХ В ДЛИТЕЛЬНОМ КОСМИЧЕСКОМ ПОЛЁТЕ // Современные проблемы науки и образования. 2008. № 5. ;URL: https://science-education.ru/ru/article/view?id=1108 (дата обращения: 06.07.2026).