



Введение
Исследованные виды Iris относятся к подроду Limniris (Tausch) Spach, секции Limniris Tausch., I. sibirica и I. sanguinea к серии Sibiricae (Diels) Lawrence, подсерии Sibiricae. I. pseudacorus к серии Laevigatae (Diels) Lawrence, I. setosa к серии Tripetalae (Diels)Lawrence [5].
Подрод Limniris, объединяющий более 45 видов, считается наиболее неоднородным по составу [3; 6]. Несмотря на то что многие виды подрода Limniris активно изучаются в систематическом отношении, репродуктивные системы остаются мало изученными или не изучены совсем.
Признаки репродуктивных органов, прежде всего данные об особенностях строения семязачатка, используются в качестве надежных критериев при выяснении дискуссионных вопросов систематики, филогении и эволюции цветковых растений. Структуры семязачатка, а также процессы их становления устойчивы, консервативны, мало зависят от изменений среды, а значит, особенно важны для целей систематики [4].
При моделировании филогенетических связей рода Iris в качестве диагностических эмбриологических признаков используются форма пыльцевых зерен, скульптура экзины, характер поверхности мембраны [2]. Структурам семязачатка же до сих пор уделяется недостаточно внимания.
Материал и методы
Объектами исследования являлись виды рода Iris: I. pseudacorus L. – и. ложноаирный, I. sanguinea Hornem. – и. кроваво-красный, I. setosa Pall. ex Link – и. щетиноносный, I. sibirica L. – и. сибирский. Исследования проведены в 2011-2013 гг. Сбор материала проводился в Пермском крае в местах естественного произрастания I. sibirica и в Ботаническом саду Ботанического института им. В. Л. Комарова РАН, за что мы выражаем благодарность куратору коллекции касатиковых Н.Б. Алексеевой.
Для изучения начальных этапов развития семязачатков фиксировались бутоны на разных стадиях развития с использованием фиксатора Кларка. Для изучения мегагаметогенеза, процесса оплодотворения, эмбрио- и эндосперматогенеза использовали метод темпоральных фиксаций, а также фиксацию завязей на разных стадиях развития после естественного опыления цветков. Фиксация завязей проводилась в фиксаторе Навашина. Проводка зафиксированного материала и парафинирование проводились по общепринятой методике. Окрашивание срезов проводилось с помощью галлоцианина – хромовых квасцов и методом тройного окрашивания [1].
Результаты
Гинецей изученных видов синкарпный, завязь нижняя. Семязачатки расположены попарно, в шахматном порядке в каждом из трех гнезд коробочки и имеют центрально-угловую плацентацию. У I. setosa отмечены единичные случаи расположения семязачатков по трое.
Мегаспорогенез у изученных видов начинается на стадии, когда ось нуцеллуса и микропиле семязачатка расположена параллельно по отношению к плаценте. Внутренний интегумент двухслойный, не выходит за пределы нуцеллуса. Наружный интегумент трехслойный, короче внутреннего. Эпидермальные клетки в основе фуникулуса радиально удлиняются, вакуолизируются, образуя фуникулярный обтуратор. Археспориальная клетка делится периклинально, отделяя париетальную клетку и мегаспороцит. Мегаспороцит характеризуется крупным ядром, плотной цитоплазмой и легко отличим от других нуцеллярных клеток. Мегаспороцит претерпевает два последующих деления мейоза, сопровождающихся цитокинезом, в результате чего образуется тетрада макроспор. Мейоз в мегаспороците I. setosa начинается одновременно с антиклинальным делением париетальной клетки.
После первого мейотического деления мегаспороцита образуются диада макроспор. У I. setosa и I. sibirica первая перегородка закладывается высоко, образуя две неравные клетки. У I. sanguinea диада состоит из равных клеток. К этому времени париетальные клетки делятся периклинально, образуя два ряда. У I. sanguinea можно наблюдать до 4 париетальных слоев. Это позволяет охарактеризовать семязачатки изученных видов как крассинуцеллятные.
После второго мейотического деления образуется линейная тетрада мегаспор. У всех изученных видов халазальная мегаспора оказывается более крупной, с большим содержанием цитоплазмы и несколько более крупным ядром, чем у остальных сестринских клеток тетрады. У I. setosa и I. sanguinea в одной завязи можно наблюдать семязачатки как на стадии мегаспороцита, так и семязачатки, в которых уже завершилось образование тетрады мегаспор после второго деления мейоза. Микропилярные мегаспоры дегенерируют и отмирают, а затем поглощаются развивающимся зародышевым мешком.
Халазальная мегаспора увеличивается, вакуолизируется и становится инициальной клеткой зародышевого мешка. Внутренний интегумент вытягивается за счет антиклинальных делений и выходит за пределы нуцеллуса. Наружный интегумент также удлиняется. Из-за неравномерного роста в области прикрепления интегументов семязачатки становятся анатропными.
После первого митотического деления образуется двуядерный ценоцит, ядра которого расходятся к полюсам зародышевого мешка. Деление ядер сопровождается образованием центральной вакуоли. Клетки париетальной ткани продолжают сдавливаться и дегенерировать (рис. 1). В результате второго митотического деления образуется четырехъядерный ценоцит (рис. 2).
Ядра остаются на полюсах и снова делятся митотически, в результате чего образуется 8-ядерный ценоцит. С прекращением митотических делений в микропилярном и халазальном концах зародышевого мешка вокруг полярных групп ядер локализуется цитоплазма и начинается клеткообразование. Период дифференциации элементов зародышевого мешка начинается сразу после клеткообразования и сопровождается интенсивным ростом зародышевого мешка.
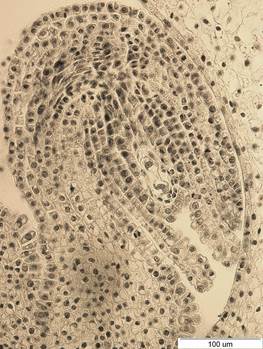
Рис. 1. Семязачаток I. sibirica на стадии двуядерного зародышевого мешка. х400.
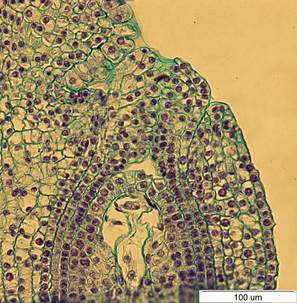
Рис. 2. Семязачаток I. pseudacorus на стадии четырехъядерного зародышевого мешка. х400.
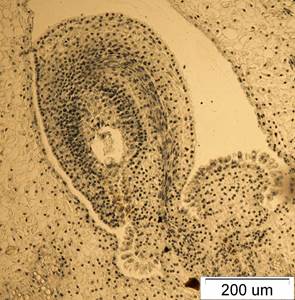
Рис. 3. Семязачаток I. sanguinea на стадии зрелого зародышевого мешка. х100.
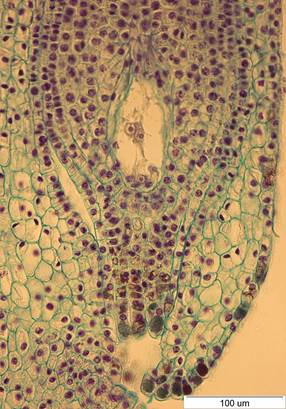
Рис. 4. Семязачаток I. setosa на стадии зрелого зародышевого мешка. х400.
У изученных видов слияние полярных ядер происходит до раскрытия цветка. У I. sanguinea и I. setosa в одной завязи наблюдаются как семязачатки на стадии сближения полярных ядер, так и семязачатки, в которых уже сформировалось вторичное ядро центральной клетки.
Клетки париетальной ткани, окружающей зародышевый мешок, сдавливаются. Клетки внутреннего интегумента продолжают делиться антиклинально. Проводящая система семяпочек хорошо развита. С ростом и развитием семязачатков проводящий пучок дифференцируется по направлению к халазе. Проводящий пучок состоит из нескольких слоев удлиненных густоплазменных клеток, часть которых затем трансформируется в сосуды с кольчатыми и спиральными утолщениями и образует участки ксилемы. У изученных видов проводящий пучок доходит до халазы (рис. 3).
Внутренний интегумент у изученных видов двухслойный, расширяется в микропилярной части. У I. pseudacorus внутренний интегумент на микропилярном конце утолщается до 3-5 слоев, у I. setosa – до 6 слоев, у I. sanguinea – до 3-5 слоев, у I. sibirica до 3 слоев. У I. sibirica имеется интегументальный обтуратор - покровные клетки, выстилающие микропиле, крупные, радиально вытянутые, с утолщенными наружными стенками и плотной цитоплазмой.
Наружный интегумент многослойный. У I. pseudacorus наружный интегумент 5-7-слойный, короче внутреннего интегумента. У I. setosa наружный интегумент 5-6-слойный, длиннее внутреннего интегумента, в области микропиле значительно расширяется до 7-10 слоев и за счет увеличения размеров клеток сужается к краю. У I. sanguinea наружный интегумент состоит из 5 слоев, по длине равен внутреннему интегументу. У I. sibirica наружный интегумент 4-6-слойный, короче внутреннего интегумента. Микропиле прямое, образовано только внутренним интегументом. Клетки обтуратора имеют утолщенные наружные стенки, плотную цитоплазму. К моменту раскрытия цветка зародышевый мешок у изученных видов состоит из яйцеклетки и двух синергид в микропилярной части, центральной клетки с вторичным ядром, трех антипод в халазальной части (рис. 4).
У I. sanguinea центральная клетка расположена вблизи антипод. В некоторых полноценно сформированных семязачатках у I. sibirica, I. sanguinea, I. setosa можно наблюдать аномальное увеличение числа ядер в недоразвитых зародышевых мешках.
Оплодотворение двойное, порогамное. Пыльцевая трубка проходит по микропиле, проникает в одну из синергид и изливает в нее содержимое. После проникновения пыльцевой трубки в зародышевый мешок синергиды начинают деформироваться. Один спермий сливается с яйцеклеткой, формируя зиготу. Другой спермий сливается с центральной клеткой (его ядро объединяется с вторичным ядром), образуя первичную клетку эндосперма. Первичная клетка эндосперма и зигота проходят короткий период покоя после двойного оплодотворения. Деление ядра первичной клетки эндосперма предшествует делению ядра зиготы.
У I. setosa и I. pseudacorus деление ядра первичной клетки эндосперма происходит на вторые сутки после опыления. Эндосперм изученных видов нуклеарный. Свободные ядра, имеющие 1-3 ядрышка, связаны между собой тяжами цитоплазмы. I. pseudacorus ядра содержат до 7 ядрышек.
Во время первого деления зиготы клеточная перегородка закладывается поперечно. У I. pseudacorus и I. setosa 2-клеточные зародыши в семязачатках можно наблюдать на шестой день после опыления. У I. pseudacorus и I. sanguinea синергиды на этой стадии полностью разрушаются. Затем в апикальной клетке закладывается продольная клеточная перегородка, в базальной клетке – поперечная. В 4-клеточном зародыше расположение клеток Т-образное. У I. setosa на этой стадии можно различить остатки дегенерирующих синергид. У I. sibirica обе синергиды сохраняются вплоть до образования многоядерного нуклеарного эндосперма. Антиподы у I. pseudacorus и I. setosa сохраняются до образования 4-клеточного зародыша. Антиподы у I. sanguinea и I. sibirica дегенерируют в начале развития эндосперма. У I. sibirica I. setosa и наблюдается увеличение числа антипод до четырех.
Таким образом, зародышевые мешки в семязачатках развиваются по Polygonum-типу. Семязачатки крассинуцеллятные, анатропные, битегмальные. В основании фуникулуса имеется область с радиально вытянутыми клетками - обтуратор – ткань секреторного типа, обеспечивающая прохождение пыльцевых трубок в зародышевый мешок.
Заключение
Анализ строения и развития семязачатков Iris pseudacorus, I. setosa, I. sanguinea и I. sibirica позволяет выделить следующие эмбриологические признаки в качестве диагностических: наличие или отсутствие радиально удлиненных клеток внутреннего интегумента, выстилающих микропиле, длина наружного интегумента относительно внутреннего, увеличение числа антипод, сохранение или дегенерация синергид и антипод после оплодотворения (табл. 1).
Таблица 1 - Эмбриологические признаки женской генеративной сферы видов рода Iris L.
|
Вид |
Внутренний интегумент |
Наружный интегумент |
Антиподы |
Синергиды |
|
Серия Sibiricae (Diels) Lawrence, подсерия Sibiricae |
||||
|
I. sibirica |
На микропилярном конце увеличивается до 3 слоев, имеется интегументальный обтуратор |
4–6-слойный, короче внутреннего |
Дегенерируют в начале развития эндосперма, увеличение до 4 |
Сохраняются до образования многоядерного нуклеарного эндосперма |
|
I. sanguinea |
На микропилярном конце увеличивается до 3 слоев |
5-слойный, равный внутреннему |
Дегенерируют в начале развития эндосперма |
Деформируются после проникновения пыльцевой трубки в зародышевый мешок |
|
Серия Tripetalae (Diels)Lawrence |
||||
|
I. setosa |
На микропилярном конце увеличивается до 6 слоев |
5–6-слойный, длиннее внутреннего интегумента |
Сохраняются до образования 4-клеточного зародыша, увеличение до 4 |
Деформируются после проникновения пыльцевой трубки в зародышевый мешок |
|
Серия Laevigatae (Diels) Lawrence |
||||
|
I. pseudacorus |
На микропилярном конце увеличивается до 3–5 слоев |
5–7-слойный, короче внутреннего |
Сохраняются до образования 4-клеточного зародыша |
Деформируются после проникновения пыльцевой трубки в зародышевый мешок |
Строение семязачатка I. sibirica отличает его от других изученных видов подрода Limniris наличием интегументального обтуратора, сохранением синергид после оплодотворения и увеличением числа антипод до 4. По мнению Макаревича с соавторами [3], I. sibirica близок к видам подрода Xyridion, а не Limniris и, по всей вероятности, наиболее близок к исходному предковому виду. Также авторы отмечают близость I. setosa с видами подрод Iris и рекомендуют включить его в этот подрод, в то время как во всех современных классификациях он помещен в подрод Limniris. У I. setosa внешний интегумент длиннее внутреннего, наблюдается увеличение числа антипод до 4 и их сохранение вплоть до начала эмбриогенеза. Указанные выше отличия в развитии женской генеративной сферы, как относительно наиболее консервативной, могут быть использованы для уточнения систематического положения и филогенетических связей видов подрода Limniris, наряду с другими таксономическими признаками.
Рецензенты:
Колясникова Н.Л., д.б.н., доцент, зав. кафедрой ботаники, генетики, физиологии растений и биотехнологий ФГБОУ ВПО «Пермская государственная сельскохозяйственная академия имени Д.Н. Прянишникова», г. Пермь.
Шамров И.И., д.б.н., доцент, зав. кафедрой ботаники РГПУ им. А.И. Герцена, г. Санкт-Петербург.
Библиографическая ссылка
Дорофеева М.М. ЭМБРИОЛОГИЧЕСКОЕ ИССЛЕДОВАНИЕ ВИДОВ IRIS L. ПОДРОДА LIMNIRIS (TAUSCH) SPACH // Современные проблемы науки и образования. 2013. № 5. ;URL: https://science-education.ru/ru/article/view?id=10746 (дата обращения: 16.06.2026).