



В радиобиологии и на сегодняшний день остается немало парадоксальных и нерешенных проблем. Например, радиобиологический парадокс воздействия низких и сверхнизких доз ЭМИ и γ-излучений, механизм стимулирующего действия природного радиационного фона, невыясненная причина возникновения асимметрии живой природы, парадокс «оживляющего» действия вторичного биогенного излучения и др. Космическая радиобиология добавила к этим парадоксам новые [22]. Среди них – вспышки в глазах космонавтов, механизм возникновения которых далек от ясности и в наши дни; неожиданное возрастание остроты зрения космонавтов на два порядка, наблюдаемые в ряде случаев [11]; загадочный механизм появления космофизических макрофлуктуаций в различных системах; эффекты локального аномального воздействия космических излучений на выживаемость и мутагенез биообъектов [5]. Есть основания предполагать, что все упомянутые парадоксы связаны с проявлением единого общего механизма, прежде всего объединяющего механизмы их слабых воздействий.
В условиях космического полета на биологические объекты действуют физические поля различной природы. В течение ряда лет авторами проводились эксперименты на биологических объектах по обнаружению локального радиационного фактора (ЛРФ) космического пространства и по измерению воздействия адронов высоких энергий (АВЭ) на протонном ускорителе по тестам выживаемости и мутагенеза на биологических объектах [18,19]. При этом у индуцированных ЛРФ мутаций исследовались молекулярные изменения, представляющие большой интерес в теории и практике космических исследований [12,16,23].
Материалы и методы: Объектами исследования служили бактериофаг Т4Вг+, семена томатов Lycopersicon esculentum Mill, конские бобы Vicia faba, самцы белых беспородных мышей. После выращивания бактериофага Т4Вг+ (БФ) на E. coli B в полноценных и синтетических питательных средах образцы БФ облучали параллельно на ускорителе и на γ-установке в стеклянных ампулах (109 – 1010 фагочастиц/мл) с толщиной стенок 1 мм и объемом 0,5 мл, а также в герметических ампулах-пробирках из оргстекла. Определение выживаемости БФ проводили методом агаровых слоев в модификации Раутенштейна с индикаторным штаммом E. coli B. Процент выживаемости подсчитывали по отношению числа выживших фагочастиц, подвергшихся облучеию, к числу исходных частиц в контроле. Высушенная в эксикаторе пластина (30% желатины + глицерин) с БФ и с 5% лактозой для лучшего сохранения выживаемости БФ использовалась для генетико-биохимических и биофизических исследований.
Семена томата длительное время облучали на орбитальном космическом комплексе «МИР», и с помощью соматических и цитогенетических тестов определяли количество аберрантных клеток, крупные и протяженные делеции. Были использованы также семена Vicia faba для определения действия γ-излучения и воздействии АВЭ при их замачивании в различных концентрациях химически веществ (10-4 – 10-5 М) в разные сроки до облучения (0,25-2 часов). По количеству проросших семян (%) определяли эффективность радиозащиты препарата во времени. Для определения протекторного индекса для других природных веществ (убихиноны, эхинохром) были использованы белые беспородные мыши весом 20-22 г., облученные в дозе 7,5 Гр на установке ГУБЭ радиоизотопом 60Со, мощность дозы 1,76 Гр/мин. Препараты, используемые для проверки модификации лучевого поражения, растворяли в этаноле или в диметилсульфоксиде и вводили внутрибрюшинно или подкожно мышам в соответствующих концентрациях за 15-60 минут до облучения. Выживаемость лабораторных мышей определяли на 30-е сутки наблюдения.
Результаты исследований и их обсуждение: В работе рассматривается воздействие светового (фотонного) и ионизирующего излучений на рецепторные молекулы родопсина с характерным размером 60Å. Структура фоторецепторных мембран наружного сегмента палочек позвоночных содержит порядка 108 – 109 молекул зрительного пигмента на сегмент и имеетвысокое его содержание в палочках сетчатки глаза (~ 85% от общего количества мембранного белка). На фрагментах фоторецепторных мембран показано, что первичные механизмы поглощения световой энергии связаны с фотоизомеризацией ретинальной компоненты родопсина из цис- в более стабильную транс-форму. Процесс сопровождался изменением конформации ретиналя и его отделением от белка опсина, а также увеличением проницаемости цитоплазматических мембран наружного сегмента палочек сетчатки для ионов Na+ [13]. Конформационные изменения белковой части родопсина сопровождались уменьшением Na+-проводимости цитоплазматических мембран. Медиатором в акте фоторецепции является цГМФ, а ионы Са+2 определяют процессы адаптации. Фосфорилирование особых белков, регулирующих проводимость Na+-каналов, зависело от циклических нуклеотидов [13]. Деполяризация мембраны в рецепторной клетке приводит к возникновению рецепторного потенциала. В окончании нейрона возникает генераторный потенциал, который при значительной величине запускает потенциал действия, ответ на энергию фотона или поляризованного света.
Хорошо известны гипотезы [21], основанные на возникновении излучения Черенкова при прохождении заряженной частицы через стекловидное тело глаза и на прямом действии ионизирующих частиц на сетчатку глаза. Для расчета вероятности вспышки по этим механизмам разработана микродозиметрическая модель [4]. Одним из авторов настоящей статьи была высказана мысль, что микродозиметрический подход, который связывает их возникновение с энерговыделением в чувствительном объеме зрительного анализатора - молекуле родопсина, мог бы быть также наиболее универсальным подходом к разработке количественной модели появления вспышек в глазах космонавтов за счет высокой чувствительности комплексов фоторецепторных дисков (в сетчатке млекопитающих содержится 1,5.1010 палочек с дисками). Флуктуации этого энерговыделения определяют вариабельность наблюдаемого эффекта и его вероятность. Для конкретного эксперимента в высокогорье с помощью микродозиметрической модели была рассчитана вероятность возникновения вспышки при прямом возбуждении родопсина ионизирующими частицами и по механизму, связанному с образованием Черенковского излучения при прохождении ионизирующей частицы через глаз. Подробное описание моделей и расчетов приведено в работе [14]. Сравнение рассчитанных вероятностей вспышки по двум механизмам с экспериментальными данными приводит к выводу, что доминирующим является механизм прямого возбуждения сетчатки. Он приводит к результатам, совпадающим по порядку величины с экспериментальными данными. Вероятность же вспышки по механизму образования Черенковского излучения при прохождении через глаз заряженной частицы оказывается на 3-4 порядка ниже по сравнению с механизмом прямого возбуждения. Однако в более поздних экспериментах на ускорителях, выполненных спещиально для определения соотношения вкладов в вероятность возникновения вспышки по двум рассматриваемым механизмам, было показано, что оба механизма приводят к примерно равным вкладам в вероятность вспышки. В связи с этим возник вопрос: «Чем объяснить столь высокую эффективность действия Черенковского излучения, несмотря на его небольшой энергетический вклад?». Требуемый коэффициент повышения эффективности воздействия Черенковского излучения для достижения согласия с экспериментом составляет 103-104. Мы обратили внимание, что это значение совпадает с коэффициентом усиления слабого воздействия. Предложенный нами критерий слабого воздействия на основе универсальной зависимости чувствительности биологической системы от возрастающего физического стимула приведен на рис. 1.
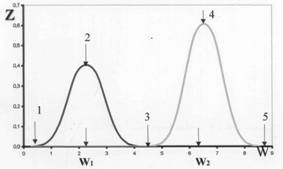
Рис. 1. Универсальная зависимость фазовой реакции биосистемы Z от возрастающего физического стимула W (мощность адекватного стимула электромагнитного излучения, концентрация микроэлемента и т.д.) в относительных единицах. По оси абсцисс – мощность физического стимула в логарифмическом масштабе, по оси ординат – фазовая реакция биосистемы. 1 - абсолютный порог чувствительности, 2 - порог включения компенсаторных систем, 3 - порог тотальной мобилизации, 4 – срыв, 5 – гибель (биологический механизм физиологической адаптации). Левый пик - «слабые» воздействия, правый пик — «сильные» воздействия (согласно физическому механизму слабых воздействий).
Анализ экспериментальных данных показывает, что отношение стимулов w1/w2 для систем различного уровня организации (от клеточного до популяционного) равно 104 (очень редко, в порядке исключения, (104)2 или (104)1/2) [6,7].
В нашем конкретном случае фиксированного пролета космического мезона мы имеем в среднем в видимой области электромагнитного спектра один Черенковский фотон, то есть нет сомнений, что это действительно слабое воздействие. Проблема слабых воздействий – актуальная общебиологическая и даже общенаучная проблема. Чернавский и Хургин, рассматривая в своем обзоре трудности ее решения, показали, что во всех предложенных механизмах слабых воздействий не хватает коэффициента усиления, не меньшего 104 [14]. Именно такой коэффициент был найден нами экспериментально [6]. Возвращаясь к нашей задаче возникновения вспышки в глазах по механизму образования Черенковского излучения, остается лишь убедиться, что Черенковское излучение поляризовано. Известно, что это действительно так [8]. С учетом этого, ранее рассчитанная вероятность вспышки от Черенковского излучения 10-4 при умножении на коэффициент усиления эффективности слабого воздействия поляризованного света (104) становится сравнимой с вероятностью вспышки при прямом возбуждении родопсина. Так разрешился парадокс «видимого» расхождения расчета и эксперимента. Аналогичным образом объясняется и другой, отмеченный выше парадокс – возрастание в ряде случаев остроты зрения космонавтов на два порядка [6]. И в этом случае все условия эффективного слабого воздействия налицо, если принять во внимание, что отраженный от земных предметов свет, попадающий в глаз космонавта на орбите, поляризован и его интенсивность мала. Учитывая, что острота зрения пропорциональна квадратному корню из интенсивности света, а возрастание эффективности воздействия эквивалентно увеличению интенсивности, то в таком случае максимально возможное увеличение остроты зрения пропорционально (104)1/2=102. Этот эффект наблюдается не всегда, т.к. поляризация света зависит от состояния атмосферы Земли и других условий. В правильности предложенного объяснения убеждает его применимость для решения и других парадоксов. Например, объяснение квантовой эффективности зрительного рецептора – палочки, равной 0,5 (а не 1) в однофотонных экспериментах без различения лево- и право поляризованных фотонов. Один из них регистрируется с эффективностью 1, другой – с близкой к 0 [6]. Обычный свет состоит из равного количества «левых» и «правых» фотонов, что и обеспечивает появление усредненного значения 0,5. Этим же объясняется парадокс высокой чувствительности глаза. Энергия, необходимая для физиологической стимуляции рецепторной клетки, в несколько тысяч раз (104) выше энергии одного фотона, ее вызывающей. Естественное объяснение получает и ряд других парадоксальных явлений: возникновение миражей (отраженный от облаков поляризованный свет); высокая чувствительность зрения «лунатиков» при хождении по карнизу (отраженный лунный свет); обнаружение поляроидных пленок на глазах глубоководных рыб и т.д. Эффект высокоэффективного воздействия поляризованного излучения был использован нами при объяснении тонкой структуры гистограмм (статистический разброс) радиоактивности и других явлений, обнаруженной Шнолем [15] и названной им космофизическими макрофлуктуациями. Ранее нами было показано, что космическим агентом, вызывающим эти макрофлуктуации, является реликтовое излучение (РИ) Вселенной, точнее, его анизотропная, поляризованная компонента, представляющая незначительную добавку к изотропной, неполяризованной компоненте (не более 10-3), но оказывающая заметное воздействие за счет высокоэффективного действия поляризованного излучения. Постоянная, изотропная компонента обеспечивает устойчивость различных систем; изменяющаяся анизотропная, поляризованная составляющая – их изменчивость. Обнаруженная нами новая, фундаментальная роль поляризованной компоненты РИ оказалась полезной при объяснении загадочной зависимости параметров физиологической системы (психопатологический синдром (ППС) в баллах) от Ар-индекса солнечной активности, установленной эмпирически Жвирблисом [7]. Характер этой связи (корреляция, антикорреляция, отсутствие корреляций) определяется секторной структурой межпланетного магнитного поля (10-4 – 10-6 Гс). Оставалось совершенно не понятным и парадоксальным влияние столь слабых магнитных полей на изменение характера связи ППС с Ар-индексом. Нам было ясно, что ответ на этот вопрос надо искать в изменении поляризованной составляющей реликта. Механизм этого изменения стал окончательно ясен, когда мы связали это изменение с эффектом Ханле – деполяризацией излучения в слабых магнитных полях, открытым в начале прошлого века. Так разрешился и этот парадокс. И, наконец, с поляризацией частиц и излучений локального ливня, образующегося при взаимодействии биообъекта с высокоэнергетической частицей космического излучения, может быть связано возможное объяснение эффектов локального воздействия космического излучения, изменяющего на порядки выживаемость и показатели мутагенеза биообъектов.
Проведенные эксперименты в космосе, высокогорье, на ускорителе и в земных условиях позволили нам связать ЛРФ с тяжелыми заряженными частицами (ТЗЧ) и АВЭ. Различие по выживаемости и генетическому действию ТЗЧ и АВЭ выявляются как на макро, так и на микро биологических уровнях. Макролокальные, крупные (очаговые) нарушения тканевых структур организмов, проявляющиеся как морфологические изменения и расплавление тканей, связывают с действием сверхжестких АВЭ космоса, регистрируемых исследователями с низкой частотой [17]. Так, на рисунке 2А представлен макролокальный эффект ЛРФ, появляющийся при действии вторичного излучения адронов (ядерных частиц) Серпуховского протонного ускорителя как модели излучения АВЭ космического пространства. ЛРФ проявлялся в виде очень низкой общей выживаемости бактериофага в двух ампулах, которая составила 0 и 102 фагочастиц/мл (при общей выживаемости в среднем 109 фагочастиц при общей средней дозе облучения 375 Гр). Рассчитанный общий эффект по совокупности всех остальных ампул должен быть равен < 10% выживаемости, тогда как в действительности в этих 2-х точках выживаемость составляла < 10-10% (различие по выживаемости в 109 раза). По перерасчетным дозам величина относительной биологической эффективности (ОБЭ) составляла > 25. В контрольном варианте при действии γ-лучей, для которых характерны в основном электромагнитные взаимодействия (рис. 2Б), выживаемость составляла 2.108 фагочастиц/мл, и подобных выбросных точек, наблюдаемых в опытном блоке ампул, никогда не наблюдалось. Отклонения выживаемости (и мутаций) бактериофага в пределах одного опыта были таковы, будто дозы облучения отдельных экспериментальных групп неожиданно изменялись в сторону повышения в 5-10 раз и более, несмотря на то, что для опытных ампул соблюдались идентичные условия облучения и были исключены возможные влияния магнитного поля, мощности дозы в отдельных экспериментальных точках. При облучении частиц в буфере отклонения достигали 7 порядков при дозе 400 Гр и 4 порядков – при дозе 800 Гр, в бульоне – 5 порядков при дозе 1200 Гр. Эти эффекты оказались незамеченными применявшимися приборами для измерения дозы излучения, которая имела неравномерное локальное распределение, но максимально отличалась в пределах одного биоблока всего в 2 раза (рис. 3). Таким образом, в отличие от γ-облучения, биологический эффект АВЭ не зависел от дозы их излучения, как в других экспериментах, проведенных на различных космических станциях и спутниках, при изучении влияния космического излучения на выход мутаций у хлореллы и Crepis capillaris [1].
Изучаемый феномен ЛРФ представлен на рис. 4, где дана уменьшенная фотография множественной генерации вторичных ядерных частиц в результате сильного взаимодействия одного иона (АВЭ) с энергией 47 ГэВ с одним ядром углерода пропановой камеры Mirabel (длина снятой области около 2 м). В начальной части узкого пучка вторичного излучения образуется локально высокая плотность ионизационных треков ТЗЧ и многозарядных фрагментов ядра и, соответственно, локально большее выделение энергии, с которыми также может быть связан высокий ЛРФ наряду с особенностями действия поляризованных частиц вторичного излучения АВЭ.
А Б
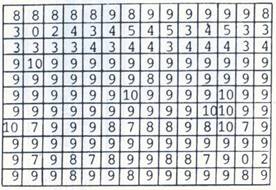
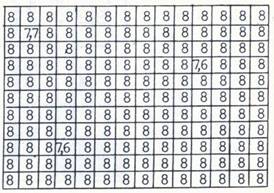
Рис. 2. Локальная выживаемость бактериофага в биоблоке при воздействии излучения ускорителя. А – в опытном биоблоке, средняя доза излучения 375 Гр, Б - контрольном биоблоке при действии γ-лучей, доза 460 Гр. Указан показатель степени количества выживших фагочастиц в 1 мл. Исходная концентрация бактериофага 5 х 1010 /мл.
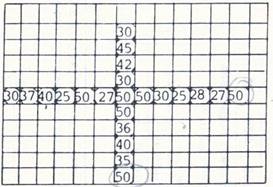
Рис. 3. Распределение локальных доз вторичного излучения от протонов 70 Гэв для биоблоков по данным термолюминесцентных и активационных детекторов, размещающихся в каждой клетке биоблока; локальные дозы находились в пределах от 250 до 500 Гр.
Последующие эксперименты по изучению микролокальных молекулярных изменений, вызванных ЛРФ космоса, высокогорья и излучений ускорителей, обнаружили, что крупных, протяженных и множественных делеций ДНК бактериофага (макромутаций) в космосе индуцировалось 65,17%, в высокогорье - 54,4 %. Подобные изменения ДНК исключали супрессорные мутации (реверсии), они обусловлены в основном ТЗЧ и в некоторой степени АВЭ [17]. Мутагенный эффект ТЗЧ, определяемый частотой мутагенеза в зонах с пролетом треков частиц, был в 10 и более раз больше, чем в необлученном контроле. При контрольном γ-облучении 137Cs, равномерно распределенном в пространстве, подобных мутаций не наблюдалось, спектр мутаций был иным. На основании наших результатов соматических и цитогенетических тестов (множественные аберрантные клетки, делеции) летального действия АВЭ были рассчитаны более низкие величины (2-5) относительной биологической эффективности (ОБЭ) для метаболизирующих биообъектов (бактерий, растений) по сравнению с ОБЭ неметаболизирующих бактериофагов (4-8), не способных к репарации in vitro[2]. Известно, что эффект γ-излучения, наоборот, выше на метаболизирующие и пролиферирующие клетки и ткани. На основании представленных данных можно сделать вывод о различном механизме действия АВЭ и γ-излучения как физических факторов космического полета (ФФКП). Суммарное влияние ФФКП по генетико-физиологическим изменениям, наблюдаемое при прорастании семян томатов, длительное время экспонированных на орбитальном комплексе «МИР» выражалось в стимуляции их прорастания, морфологически выраженных генетических мутациях [12].
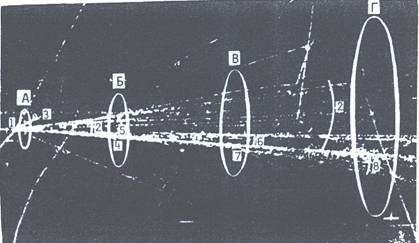
Рис. 4. Результат взаимодействия одного иона с энергией 47 ГэВ с ядром углерода в пропановой камере Серпуховского протонного ускорителя Института физики высоких энергий
Обозначения: 1 – первичное сильное взаимодействие; 2 – узкий пучок вторичных заряженных ядерных частиц; 3 – след многозарядного фрагмента (ТЗЧ); 4,7 – вторичные сильные взаимодействия вторичных адронов; 5 – след ТЗЧ, рожденных вторичным взаимодействием; 6,8 – узкие пучки ядерных частиц, генерированных адронами пучка вторичных частиц. Проявления в виде вспышек: А – ярких, малых, Б – диффузионного «облака»; В – двойное «облако», Г – полосы типа «снега» при окончательном распаде каскада на γ-кванты, электроны, нейтроны и другие вторичные частицы.
Защита от АВЭ при работе на ускорителях и в космических полетах имеет большое значение в связи с их высокой локальной биологической активностью. Поиск эффективных средств защиты, однако, затруднителен, поскольку биообъекты обладают неэффективными репарационными системами и эволюционно не приспособлены к воздействию этого фактора в земных условиях. Задача радиозащиты от АВЭ осложняется также большой вариабельностью результатов при близких условиях экспериментов, связанной с крайней неравномерностью распределения поглощенной энергии в микро- и макрообъемах. Нами был получен более высокий антимутагенный радиозащитный эффект от γ-излучения, чем от АВЭ в дозе 0,2-10 Гр, полученных на ускорителе, при выращивании бактерий E.coli и бактериофага Т4 в пептонном бульоне. Действие антиоксидантов также оказалось более эффективным при защите от γ-излучения, чем от АВЭ, что связано с макролокальным и высокоэффективным действием АВЭ на биологические объекты. Замачивание семян Vicia faba в течение 0,25-2 часов в 10-4 – 10-5 М растворе дибунола или эхинохрома с последующим облучением АВЭ приводило к увеличению эффективности их прорастания на 10-15%. При γ-облучении лабораторных мышей в летальной дозе 7,5 Гр дибунол в тех же концентрациях на 60-70% увеличивал выживаемость животных, а убихиноны Q-6, Q-9, Q-10 в ДМСО (10-2 М), введенные за час до облучения, - на 80-90% [10]. Был показан высокий профилактический и терапевтический эффект также и для других природных нетоксических антиоксидантов при действии сублетальных и летальных доз γ-излучения 5-8 Гр – эхинохрома, β-каротина, флавонолов, антоцианов и дигидрокверцитина, взятых в концентрации 10-6 – 10-3 М. Первичные механизмы их действия на молекулярно-клеточном уровне реализуются в их способности связывать свободные радикалы, а также изменять протонную проводимость биомембран, стимулировать синтез макроэргов (АТФ) в мембранах митохондрий, регулировать транспорт ионов Са2+, К+, Na+, Cl-, изменять потенциал и структуру липидной части мембран [9]. Эти мебранотропные препараты в низких концентрациях обладали пролонгированностью действия, высоким протекторным индексом и низкой токсичностью, они могут быть использованы в условиях космических полетов при более низких дозах γ-излучения с более высокой эффективностью их радиозашитного действия. Низкоинтенсивное РИ, может, наоборот, использоваться для усиления адаптационных способностей человека и животных к изменяющимся условиям окружающей среды и в лечебных целях [9]. Так, было показано снижение содержания ретикулоцитов, повышенного в периферической крови крыс после кровопотери, что связано с замещением анаэробного энергообмена аэробным, обусловленным депротонированием ферментов, участвующих в синтезе АТФ [3]. Восстановление количества лизосом в перитонеальных клетках моноцитарного и нейтрофильного рядов свидетельствовало об ускорении репарации защитных функций этих клеток [3]. Приведенные данные указывают на адаптационные свойства РИ. В настоящее время, однако, его управляющая роль заметно ослабла из-за бурного развития радио- и телекоммуникационных систем на частотах РИ, поэтому целесообразно применение в физиотерапевтических и лечебных целях моделированного ЭМИ, близкого по параметрам к РИ.
Заключение: В работе приводятся результаты исследований по воздействию ФФКП на биологические объекты. Основным ФФКП является ионизирующая радиация, сопровождающая γ-излучение, действие ТЗЧ и АВЭ, и приводящая к ионизации молекул и атомов, образованию свободных радикалов, активных форм кислорода, гидроперекисей и различных токсинов. Предлагаются эффективные мембранотропные средства защиты, обладающие антиоксидантным действием и стимулирующие синтез АТФ, испытанные на лабораторных беспородных мышах. Предложен новый механизм и модель возникновения вспышек в глазах космонавтов (и объяснения других парадоксальных явлений) на основе действия на глаз космонавтов ионизирующего космического излучения АВЭ и ТЗЧ и высокоэффективного низкоинтенсивного поляризованного света, исходящего от атмосферы Земли.
Получение нами АВЭ и ТЗЧ на Серпуховском протонном ускорителе при взаимодействии высокоэнергетического иона (АВЭ) с ядром углерода сопровождалось множественностью генерации вторичных заряженных и незаряженных частиц с их узким угловым распределением в пучке. Эти ядерные частицы обладали локальным характером воздействия. АВЭ при взаимодействии с биологическим объектом вызывали редкие, но сильные макролокальные повреждения на уровне тканей и целых организмов, например, гибель клеток бактериофага Т4Br+ в наших экспериментах. ТЗЧ космоса довольно часто индуцируют значительные микролокальные повреждения в виде специфических макромутаций (протяженных и множественных делеций ДНК) и множественных повреждений микроструктур (мембран, субклеточных частиц). Как следует из наших более ранних данных, именно такие макромутации и являются тем фактором, который обеспечивает кардинальную изменчивость генома, необходимую для эволюционного развития биологических видов. Эксперименты и расчеты показали, что ТЗЧ космического пространства ответственны в основном за микролокальные изменения клеточных структур, а АВЭ – за крупные макролокальные изменения тканей и организмов. Для защиты от γ-излучений АВЭ и ТЗЧ предлагается ряд природных нетоксичных мембранотропных соединений с антиоксидантными и энергостимулирующими свойствами.
Низкоинтенсивное РИ, исходящее из центра Вселенной, может благотворно воздействовать на живые организмы своими двумя составляющими: поляризованной и неполяризованной, поскольку являлось, по всей вероятности, мощным эволюционным фактором зарождения, адаптации и развития жизни на Земле.
СПИСОК ЛИТЕРАТУРЫ:
1. Акоев И.Г., Дубинин Н.П., Юров С.С., Ваулина Э.Н., Костина Л.Н., Ваулин Е.П. // Успехи космической биофизики. Пущино. 1978. С. 6-21.
2. Акоев И.Г., Юров С.С. // Проблемы космической биологии. 1989. Т. 60. С. 60-80.
3. Даровских С.Н., Рассохин А.Г., Кузнецов М.Е. // Биомедицинские технологии и радиоэлектроника. 2005. № 6. С. 40-46.
4. Дмитриевский И.М., Гас Д.Б., Щеглов В.С., Aродзеро A.И., Граменицкий П. В., Фетисов И.К. // Проблемы биологического действия и дозиметрии тяжелых заряженных частиц и адронов высоких энергий ускорителей и космического пространства. // Всес. конф. Пущино. 1984. С. 187-193.
5. Дмитриевский И.М. // Сб.: «Стратегия и тактика жизни в условиях планетарного экологического кризиса» в 3 тт. С-Пб. Изд. Гуманистика. 2002. Т. 1. С. 174-183.
6. Дмитриевский И.М. // Там же. Т. 2. С. 268-276.
7. Жвирблис В.Е. // Космофизические источники асимметрии живых систем. Принципы симметрии и систематики в химии. Под ред. Степанова Н.Ф. Изд. МГУ. М. 1987. С. 87-106.
8. Жевандров Н.Д. // Анизотропия и оптика. Москва. Наука. 1974. С. 31-34.
9. Кожокару А.Ф. // Автореф. дисс. на соискание уч. степ. докт. физ.-мат. наук. Направленная модификация структурно-функционального состояния мембран с целью изменения устойчивости клеток и организма к факторам окружающей среды. М., 1992. 88 с.
10. Кожокару А.Ф., Заславский Ю.А., Алексеева Л.В., Акоев И.Г. // Радиобиология. 1981. Т. 22. № 5. С. 693-696.
11. Лазарев А.В., Николаев А.Г., Хрунов Е.В. // Оптические исследования в космосе. Гидрометеоиздат. Ленинград. 1983. С. 121-123.
12. Нечитайло Г.С., Юров С.С., Кожокару А.Ф., Ревин А.Ф. // XV Российский национальный конгресс «Человек и лекарство». Москва. 14-18 апреля 2008. С. 674-675.
13. Фесенко Е.Е. Первичные процессы рецепции запаха и света. Автореф. дисс. на соискание уч. степ. докт.биол.наук. Пущино. 1980. 48 с.
14. Чернавский Д.С., Хургин Ю.И. // Миллиметровые волны в медицине и биологии. Под ред. Девяткова Н.Д. Москва. 1989. С. 227-235.
15. Шноль С.Э., Коломбет В.А. и др. УФН. 1998. Т. 168. № 10. С. 1129-1140.
16. Юров С.С., Кожокару А.Ф., Дмитревский И.М. // Вестник Российской военно-медицинской академии. 2008. № 3 (23), приложение 1. С. 227.
17. Юров С.С., Ливанова И.А., Смирнова Е.Н. и др. // Успехи космической биофизики. Пущино. 1978. С. 38-44.
18. Akoev, I.G., S.S. Yurov. // Life Science and Space Research. X111-Akademie-Verlag, Berlin. 1975. P. 187-193.
19. Akoev, I.G., Yurov S.S., Akoev V.I. // Adv. Space Res. 1981. Vol. 1. Р. 75-81.
20. Dmitrievskiy, I.M. // Biophysics, 2001. Vol. 46, 852-853.
21. Fasio, G.G., Jolley I. V., W.N Charman. // Nature. 1970. Vol. 228. P. 260-261.
22. Nechitailo, G. S., and A.L. Mashinsky. // Space biology. Studies at orbital stations. ”Mir”. Moscow. 1993. P. 362-365.
23. Mashinsky, A.L., G.S. Nechitailo. // Space Res. 2001. Vol. 39. 4, P. 1-12.
Библиографическая ссылка
Дмитриевский И.М, Юров С.С, Кожокару А.Ф, Нечитайло Г.С МЕХАНИЗМЫ ДЕЙСТВИЯ ИОНИЗИРУЮЩИХ ИЗЛУЧЕНИЙ ПРИ ВОЗНИКНОВЕНИИ ЛОКАЛЬНЫХ ИЗМЕНЕНИЙ БИОЛОГИЧЕСКИХ КЛЕТОЧНЫХ СТРУКТУР В УСЛОВИЯХ КОСМИЧЕСКОГО ПОЛЕТА // Современные проблемы науки и образования. 2008. № 5. ;URL: https://science-education.ru/ru/article/view?id=1045 (дата обращения: 13.07.2026).