


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
CHANGES IN THE CONTENT OF EPIDERMAL GROWTH FACTOR AND THE SOLUBLE FORM OF ITS RECEPTOR DURING THE ANTIMETASTATIC ACTION OF A NEW EGFR INTERNAL DOMAIN BLOCKER, A DERIVATIVE OF PYRIMIDIN-4(1H)-ONE

Введение
Применение в онкологии таргетных терапевтических подходов с использованием низкомолекулярных ингибиторов, моноклональных антител, иммунотерапии, конъюгатов «антитело – лекарственный препарат» позволило значительно повысить эффективность терапии ряда злокачественных новообразований и уменьшить системную токсичность противоопухолевого лечения. Ингибиторы тирозинкиназы (ИТК) стали применяться в терапии как гематологических злокачественных новообразований, так и солидных опухолей (хронический миелоидный лейкоз, немелкоклеточный рак легкого (НМРЛ), HER2-положительный рак молочной железы и пр.). Однако обратной стороной таргетной терапии является развитие специфической токсичности и неизбежной резистентности опухолей, обусловленной как генетическими, так и негенетическими механизмами. Их изучение и разработка новых молекул, позволяющих преодолеть резистентность (первичную, вторичную), сохраняет свою актуальность еще и в связи с тем, что применение этого метода существенно ограничено при лечении опухолевой болезни на поздних стадиях.
Широко применяемыми в настоящий момент являются низкомолекулярные ИТК рецептора эпидермального фактора роста (EGFR), конкурирующие с АТФ за связывание с внутриклеточным тирозинкиназным доменом белка ErbB, нарушая фосфорилирование и последующую передачу сигналов путей фосфатидилинозитол-3-киназы/Akt (PKB), Ras/Raf/MEK/ERK1/2 и фосфолипазы C (PLCγ). Для лечения рака были разработаны ингибиторы нескольких поколений, включая эрлотиниб, гефитиниб, афатиниб, осимертиниб и пр. (всего 16), показавшие клиническую эффективность только при нескольких типах рака – химиорезистентный НМРЛ, HER2+ рак молочной железы, медуллярный рак щитовидной железы, колоректальный рак, рак носоглотки, глиомы. Однако исследования показывают их эффективность и при лечении меланомы, развивающейся на слизистых [1]. Терапия этой опухоли все еще остается серьезной проблемой в связи со значительной гетерогенностью меланомы и ее зависимостью от множества факторов, в том числе генетических, фенотипических и экологических. Анализ транскрипционного профиля меланомы выявил три подтипа опухоли с разными биологическими программами, при этом более агрессивный фенотип характеризовался усиленной активацией проопухолевых сигналов в эпидермисе и тесным взаимодействием эпителия и меланоцитов, что может способствовать инвазии и метастазированию опухоли [2]. Исследования, проведенные на моделях in vivo и ex vivo, показали тесную связь прогрессирования меланомы с эпидермальной сигнализацией и дифференцировкой кератиноцитов [3]. Активация сигнального пути EGFR выявлена при изучении причин устойчивости меланомы к антиBRAF-терапии [4]. Результаты приведенных исследований свидетельствуют о необходимости изучения возможности воздействия на эту мишень – EGFR.
В Пятигорском медико-фармацевтическом институте методом молекулярной динамики было проведено моделирование ряда соединений производных пиримидина, обладающих способностью блокировать внутренний тирозинкиназный домен EGFR, разработан и произведен их синтез, определены константы ингибирования, проведены фармакологические исследования, а также подтверждено наличие противоопухолевого эффекта разной степени выраженности, имеющего половую специфичность действия в отношении меланомы В16 [5]. Это связано с фундаментальной ролью половых различий во всех аспектах онкопатологии, разной чувствительностью к гормональной стимуляции эстрогенами опухолей, развившихся в мужском и женском организмах, а также активацией эстрогенами сигнальных путей, кроме пути EGF, которые могут быть задействованы в опухолевом росте при «выключении» пути EGF [6].
Еще одной существенной особенностью соединений, разработанных и синтезированных в Пятигорском медико-фармацевтическом институте, является их способность проявлять свою активность при парентеральном способе введения в отличие от применяемых в настоящее время препаратов из данной группы низкомолекулярных ингибиторов EGFR, которые вводятся в организм перорально. Такая особенность отечественных субстанций имеет преимущество возможного использования у онкологических больных с коморбидными заболеваниями желудочно-кишечного тракта, ограничивающими всасывание уже применяемых таргетных препаратов.
Цель исследования – сравнительное изучение противоопухолевой активности новой отечественной субстанции, производной пиримидина, и изменения под ее влиянием содержания эпидермального фактора роста (EGF) и растворимой формы его рецептора (sEGFR) в опухоли, перифокальной зоне, подмышечных и паховых лимфоузлах у самок мышей линии C57BL/6 с меланомой В16.
Материал и методы исследования
В Пятигорском медико-фармацевтическом институте было проведено молекулярное конструирование в ряду производных пиримидин-4(1Н)-она, позволившее предположить выраженную противоопухолевую активность, направленную на блокаду внутреннего домена EGFR, в сочетании с низкой токсичностью соединения 4-{2-[2-(4-гидрокси-3-метоксифенил)-винил]-6-этил-4-оксо-5-фенил-4H-пиримидин-1-ил}-бензсульфамида (6EPSVan) [7]. Для улучшения растворимости в воде его переводили в натриевую соль 6EPSVan-Na (рисунок).
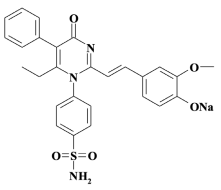
Структурная формула водорастворимой натриевой соли 4-{2-[2-(4-гидрокси-3-метоксифенил)-винил]-6-этил-4-оксо-5-фенил-4H-пиримидин-1-ил}-бензсульфамида (6EPSVan-Na)
В качестве препарата сравнения использовали гефитиниб в дозе, рассчитанной исходя из рекомендованной дозы для людей в пересчете на мышь.
Изучение противоопухолевой активности соединения 6EPSVan-Na проводили на модели in vivo – были использованы самки мышей линии C57BL/6 массой 25–28 г (n=40), полученные из филиала «Андреевка» ФГБУН «Научный центр биомедицинских технологий ФМБА России» (Московская область). Животных содержали в стандартных условиях вивария при естественном освещении, со свободным доступом к воде и пище. Эксперимент проводили согласно Директиве 2010/63/ЕU «О защите животных, используемых для научных целей», «Международным рекомендациям по проведению медико-биологических исследований с использованием животных». Исследование было одобрено Комиссией по биоэтике ФГБУ «НМИЦ онкологии» Минздрава России (протокол №1/229 от 21.04.2025).
Для перевивки использовали сингенный штамм меланомы В16/F10, полученный из НМИЦ онкологии им. Н.Н. Блохина (Москва). Опухоль перевивали 30 мышам подкожно в подлопаточную область справа в объеме 0,3 мл 25%-й суспензии жизнеспособных клеток меланомы, приготовленной в 0,9% растворе натрия хлорида (2-й пассаж in vivo). Жизнеспособность клеток перед перевивкой определяли с помощью теста на двойное окрашивание акридиновым оранжевым и DAPI на флуоресцентном анализаторе ADAMII LS (Nano Entek, Корея). После перевивки животных случайным образом распределили на три группы – контрольная, сравнения, основная (n=10 в каждой). Помимо этого 10 мышей включили в интактную группу. Начиная со дня перевивки всем животным проводили экспериментальную терапию ежедневно утром в течение 5 дней, затем 2 дня перерыв, затем схему повторили 1 раз. В таком виде экспериментальная терапия моделирует раннее профилактически-терапевтическое воздействие, а не лечение сформированной опухоли. В контрольной группе мышам внутримышечно вводили 0,1 мл воды для инъекций; в группе сравнения перорально через зонд вводили 0,1 мл водного раствора гефитиниба в дозе препарата 43,4 мг/кг массы мыши; в основной группе внутримышечно вводили 0,1 мл водного раствора изучаемого вещества 6EPSVan-Na в дозе 15,0 мг/кг массы мыши (разовая доза 0,375 мг/мышь в сутки), растворенных ex tempore в воде для инъекций. С 13-х суток от перевивки опухоли терапию прекратили и за животными вели ежедневное наблюдение с оценкой 2 раза в неделю динамики массы животных и объема подкожных опухолей. Объем опухоли вычисляли по формуле для эллипсоида: а * b * c * 0,52, где a, b, c – размеры опухоли, измеренные штангенциркулем в трех взаимно перпендикулярных направлениях. На 18-е сутки со дня перевивки всех мышей подвергли эвтаназии (декапитация). При некропсии отмечали наличие макрометастазов, лимфоузлы с измененной пигментацией считали метастатически пораженными.
Гистологическое исследование лимфоузлов проводили с использованием стандартных методов (фиксация образцов в 10% формалине, заливка в парафиновые блоки, окраска срезов толщиной 5–7мкм гематоксилином и эозином). Микроскопию препаратов осуществляли на микроскопе Leica DMLS2 с цифровой камерой OLYMPUS Camedia C-5050.
Из тканей опухоли (ОП), перифокальной зоны (ПЗ), кожи, не пораженной опухолевым процессом, а также подмышечных и паховых лимфоузлов (ЛУ) готовили 10% цитозольные фракции на 0,1М калий-фосфатном буфере рН 7,4, содержащем 0,1% Твин-20 и 1% БСА. Методом ИФА определяли содержание EGF (RayBio, США) и sEGFR (R&D systems, США).
Статистическую обработку данных проводили с использованием пакета статистических программ для медицинских исследований. Соответствие распределения нормальному оценивали с помощью критерия Шапиро – Уилка. Результаты представлены в таблицах в виде среднего значения±стандартная ошибка среднего (M±m). Для оценки значимости различий долей использовали точный критерий Фишера. Значимость различий объема опухолей, содержания EGF и sEGFR оценивали с помощью U-критерия Манна – Уитни и t-теста Стьюдента (в зависимости от вида распределения). Различия считали значимыми при попарных сравнениях – при р≤0,05, при множественных сравнениях – при р≤0,017 (применялась поправка Бонферрони).
Результаты исследования и их обсуждение
В контрольной группе опухоль вышла на 3-и сутки после перевивки у 9 (90%) мышей, на 5-е сутки опухоль пальпировалась у 100% животных. В группах, получавших гефитиниб и 6EPSVan-Na, на 3-и сутки после перевивки опухоль пальпировалась у 1 (10%) мыши (р=0,0011, относительно контрольной группы) и 2 (20%) (р=0,0055, относительно контрольной группы) мышей, соответственно, на 6-е сутки – у 100% мышей. После окончания экспериментальной терапии на 6-е сутки (18-е сутки с момента перевивки) средний объем опухоли в контрольной группе составил 4,03±1,24 см3, а в группах с терапией – 1,36±0,28 см3 в группе с гефитинибом (р=0,0147, по сравнению с контролем), 2,50±0,53 см3 в группе с 6EPSVan-Na (р=0,1230, по сравнению с группой сравнения). Таким образом, средний объем опухоли в группах с терапией был на 30–60% меньше, чем в контрольной группе. При некропсии в основной группе были отмечены значительные некротические изменения ткани опухоли, составлявшие более 50% ее объема. Метастатическое поражение подмышечных ЛУ наблюдалось у 100% мышей контрольной группы, 6 (60%) мышей группы сравнения (р=0,0867, по сравнению с контролем) и у 4 (40%) мышей основной группы (р=0,0108, по сравнению с контролем). Паховые ЛУ были поражены у 100% животных контрольной и группы сравнения, в основной группе – у 7 (70%) мышей, что значимо не отличалось от контроля (р=0,2105). Синхронное поражение ЛУ обеих анатомических зон отмечено у 100% мышей контрольной группы, у 6 (60%) мышей в группе сравнения (p=0,0867, по сравнению с контролем) и только у 3 (30%) мышей в основной группе (р=0,0031, по сравнению с контролем). Кроме этого, у мышей с экспериментальной терапией отсутствовали макрометастазы в легких (р=0,0007, по сравнению с контролем) в отличие от контрольной группы, в которой у 8 (80%) мышей обнаружено метастатическое поражение легких. Таким образом, выявленные в условиях данного эксперимента статистически значимые различия с контрольной группой и отсутствие таких различий с группой с гефитинибом по ряду показателей свидетельствуют о том, что новое вещество 6EPSVan-Na в целом обладает противоопухолевой активностью, сопоставимой с активностью гефитиниба, а также более выраженным антиметастатическим эффектом.
Проведенное гистологическое исследование показало наличие тотального метастатического поражения клетками меланомы ЛУ у мышей контрольной группы. После воздействия 6EPSVan-Na в корковом слое и мякотных тяжах ЛУ, пораженных опухолью, сохранялись небольшие участки скопления опухолевых клеток с признаками деструктивных изменений ядра и цитоплазмы, активацией соединительнотканных элементов, формированием комплексов лимфоцитов и плазмоцитов. После воздействия гефитинибом микрокартина ЛУ с метастатическим ростом опухоли демонстрировала иные признаки гибели клеток меланомы (фрагментация ядра, конденсирование и распыление хроматина, апоптотические тельца), что было характерно для апоптоза. Таким образом, результаты гистологического исследования дали основание предположить различный механизм реализации гибели опухолевых клеток: гефитиниб – апоптоз, 6EPSVan-Na – мобилизация клеточных и тканевых элементов защиты.
Результаты изучения содержания EGF и sEGFR в образцах опухоли, ее перифокальной зоны и кожи, не пораженной опухолевым процессом, при проведении экспериментальной терапии 6EPSVan-Na и гефитинибом представлены в табл.1. Было обнаружено, что в контрольной группе содержание EGF в опухоли было выше в 2,2 раза, в перифокальной зоне– выше в 21,0 раза, в коже, не пораженной опухолевым процессом, – выше в 4,3 раза, чем показатель в коже интактных мышей. При этом содержание sEGFR в опухоли было выше в 2,5 раза, в перифокальной зоне – выше в 2,4 раза, в коже, не пораженной опухолевым процессом,– выше в 1,6 раза, чем показатель в коже интактных мышей. В результате таких изменений соотношение sEGFR / EGF в ткани опухоли не отличалось от величины в коже интактных мышей, но в перифокальной зоне и в коже, не пораженной опухолевым процессом, было ниже соответственно в 8,8 и в 2,9 раза.
Таблица1
Содержание EGF и sEGFR в образцах опухоли, ее перифокальной зоны и кожи, не пораженной опухолевым процессом, при экспериментальной терапии 6EPSVan-Na и гефитинибом
|
Образец |
EGF, пг/г ткани |
sEGFR, нг/г ткани |
sEGFR / EGF, у.е. |
|
Кожа интактных мышей |
1,7±0,21 |
46,0±5,0 |
31744,4±6709,4 |
|
Опухоль |
|||
|
Контрольная группа |
3,8±0,42 p1=0,0002 |
114,9±11,6 p1=0,0000 |
36031,6±6262,1 |
|
Группа сравнения |
6,4±0,58 p1=0,0000 p2=0,0019 |
76,6±9,3 p1=0,0098 p2=0,0192 |
13180,5±2082,3 p1=0,0165 p2=0,0028 |
|
Основная группа |
2,0±0,19 p2=0,0009 p3=0,0000 |
62,2±6,4 p2=0,0008 |
33169,4±4435,0 p3=0,0007 |
|
Перифокальная зона опухоли |
|||
|
Контрольная группа |
35,7±3,7 p1=0,0000 |
110,9±11,3 p1=0,0000 |
3616,2±721,5 p1=0,0006 |
|
Группа сравнения |
4,2±0,42 p1=0,0000 p2=0,0000 |
19,8±1,9 p1=0,0001 p2=0,0000 |
5442,9±916,0 p1=0,0011 |
|
Основная группа |
3,1±0,33 p1=0,0022 p2=0,0000 |
48,5±5,2 p2=0,0000 p3=0,0000 |
16930,0±2216,8 p2=0,0003 p3=0,0011 |
|
Кожа, не пораженная опухолевым процессом |
|||
|
Контрольная группа |
7,3±0,73 p1=0,0000 |
73,2±7,1 p1=0,0057 |
10931,7±1298,5 p1=0,0070 |
|
Группа сравнения |
2,7±0,28 p1=0,0095 p2=0,0000 |
53,1±4,4 p2=0,0266 |
23219,3±4523,4 p2=0,0126 |
|
Основная группа |
2,4±0,26 p2=0,0000 |
52,6±5,5 p2=0,0337 |
25143,3±4354,0 p2=0,0058 |
Примечание: данные представлены в виде среднего значения±стандартная ошибка (M±m). Статистически значимые различия: р1 – по сравнению с показателем в коже интактных мышей, р2 – по сравнению с показателем в контрольной группе, р3 – по сравнению с показателем в группе сравнения. Критический уровень значимости при множественных сравнениях р=0,017 (с поправкой Бонферрони), в остальных случаях р=0,05.
Составлена авторами на основе полученных данных в ходе исследования.
В группе сравнения (экспериментальная терапия гефитинибом) содержание EGF в опухоли было выше в 1,7 раза, чем соответствующий показатель в контрольной группе, а в перифокальной зоне и в коже, не пораженной опухолевым процессом, – ниже в 8,5 и в 2,7 раза соответственно, чем в аналогичных образцах в контрольной группе. Содержание sEGFR в опухоли было в 1,9 раза ниже, чем в опухоли в контрольной группе, но в 1,7 раза выше, чем показатель в коже интактных мышей. При этом содержание sEGFR в перифокальной зоне и в коже, не пораженной опухолевым процессом, было ниже в 5,6 и в 1,4 раза соответственно, чем в контроле. Величина соотношения sEGFR/EGF в опухоли была статистически значимо ниже в 2,7 раза, чем в контроле, в перифокальной зоне не отличалась от контроля, а в коже, не пораженной опухолевым процессом, была статистически значимо выше (в 2,1 раза), чем в контроле (табл.1).
В основной группе (экспериментальная терапия 6EPSVan-Na) в образцах всех тканей было более низкое содержание EGF по сравнению с контрольной группой – в опухоли в 1,9 раза, в перифокальной зоне в 11,5 раза, в коже, не пораженной опухолевым процессом, в 3,0 раза. Содержание sEGFR также было более низким, чем в контроле, – в опухоли и в коже, не пораженной опухолевым процессом, в среднем в 1,6 раза, а в перифокальной зоне ниже 2,3 раза. Величина соотношения sEGFR/EGF в опухоли не отличалась от уровня в контрольной группе, в то же время в перифокальной зоне и в коже, не пораженной опухолевым процессом, она была выше в 4,7 и 2,3 раза соответственно. Важно отметить, что у мышей основной группы содержание EGF и sEGFR в опухоли было статистически значимо ниже в 3,2 и 1,6 раза, чем у мышей группы сравнения. В перифокальной зоне содержание EGF статистически значимо не отличалось, sEGFR было в 2,4 раза выше аналогичных показателей у мышей группы сравнения. При этом величина соотношения sEGFR/EGF в основной группе в опухоли и перифокальной зоне была в среднем в 3,1 раза выше, чем в аналогичных образцах в группе сравнения (табл.1).
Особый интерес представляло изучение содержание EGF и sEGFR в подмышечных и паховых лимфоузлах (ЛУ) у животных при проведении экспериментальной терапии, результаты представлены в табл.2. Установлено, что в контрольной группе содержание EGF и sEGFR в подмышечных ЛУ было выше в среднем в 2,5 раза, а в паховых – выше в 4,7 и 3,4 раза соответственно, чем у интактных мышей. Однако соотношение sEGFR/EGF не отличалось от величины у интактных животных в ЛУ обеих локализаций, что говорит о сохранении баланса при изменении содержания EGF и sEGFR при росте опухоли без воздействия на нее химиопрепаратов (табл.2).
Таблица2
Содержание EGF и sEGFR в образцах лимфатических узлов при экспериментальной терапии 6EPSVan-Na и гефитинибом
|
Образец |
EGF, пг/г ткани |
sEGFR, нг/г ткани |
sEGFR / EGF, у.е. |
|
Подмышечные лимфоузлы |
|||
|
Интактные мыши |
2,3±0,25 |
7,9±0,83 |
3842,4±550,3 |
|
Контрольная группа |
6,1±0,62 p1=0,0000 |
18,3±1,97 p1=0,0001 |
3241,1±504,6 |
|
Группа сравнения |
1,9±0,19 p2=0,0000 |
10,2±1,0 p2=0,0018 |
6072,7±1079,3 p2=0,0288 |
|
Основная группа |
1,3±0,13 p2=0,0000 p3=0,0187 |
7,3±0,79 p2=0,0000 |
6349,4±986,2 p1=0,0395 p2=0,0117 |
|
Паховые лимфоузлы |
|||
|
Интактные мыши |
2,1±0,19 |
9,7±0,93 |
5356,5±993,4 |
|
Контрольная группа |
9,9±1,0 p1=0,0000 |
33,2±3,8 p1=0,0000 |
3934,8±853,3 |
|
Группа сравнения |
3,6±0,37 p1=0,0021 p2=0,0000 |
14,0±1,4 p2=0,0001 |
4272,7±536,3 |
|
Основная группа |
1,7±0,18 p2=0,0000 p3=0,0002 |
13,5±1,5 p2=0,0001 |
9741,3±2017,6 p2=0,0163 p3=0,0174 |
Примечание: данные представлены в виде среднего значения±стандартная ошибка (M±m). Статистически значимые различия: р1 – по сравнению с показателем в лимфоузлах интактных мышей, р2 – по сравнению с показателем в контрольной группе, р3 – по сравнению с показателем в группе сравнения. Критический уровень значимости при множественных сравнениях р=0,017 (с поправкой Бонферрони), в остальных случаях р=0,05.
Составлена авторами на основе полученных данных в ходе исследования
Экспериментальная терапия в обеих группах приводила к снижению содержания EGF и sEGFR как в подмышечных, так и в паховых ЛУ. Так, в группе сравнения содержание EGF было ниже в 3,2 раза в подмышечных ЛУ и в 2,8 раза в паховых ЛУ, содержание sEGFR также было ниже в 1,8 и 2,4 раза соответственно в подмышечных и паховых ЛУ, по сравнению с показателями в контроле. При этом величина соотношения sEGFR/EGF была выше в 1,9 раза только в подмышечных ЛУ (табл.2).
В основной группе содержание EGF было ниже в 4,7 и 5,8 раза соответственно в подмышечных и паховых ЛУ, по сравнению с уровнем в контроле; при этом при воздействии 6EPSVan-Na этот показатель оказался в среднем в 1,8 раза более низким в ЛУ обеих локализаций, чем в группе сравнения. Содержание sEGFR также было ниже в среднем в 2,4 раза в ЛУ обеих локализаций, чем в контроле. В результате таких изменений величина соотношения sEGFR/EGF была выше в среднем в 2,2 раза в обеих группах ЛУ по сравнению с контролем, при этом в паховых ЛУ она была выше в 2,3 раза, чем у мышей с введением гефитиниба (табл.2).
При анализе полученных результатов можно отметить, что у мышей контрольной группы (рост опухоли без воздействия препаратами) содержание EGF было закономерно более высокое как в опухоли, так и гораздо более выраженно – в перифокальной зоне, а также в подмышечных и паховых ЛУ, которые были метастатически поражены у всех мышей контрольной группы. Такой результат согласуется с ролью этого фактора роста в опухолевом процессе – участие в пролиферации, выживании опухолевых клеток и метастазировании посредством аутокринной/паракринной регуляции активности EGFR и нижележащих внутриклеточных сигнальных путей, а также в индукции процессов опухолевого ангиогенеза и лимфангиогенеза за счет гиперэкспрессии сосудисто-эндотелиального фактора роста (VEGF). Одним из механизмов усиления активности EGFR-зависимых сигнальных путей в опухолях является увеличенная продукция его лиганда EGF [8]. Зависимость от сигнальных путей EGFR наблюдается не только в опухолях эпителиальной природы (рак легкого, яичников, толстой кишки и пр.), но показана и в меланомах, что обусловлено большой гетерогенностью этого типа неоплазий по экспрессируемым генам и возможной связью агрессивных фенотипических проявлений меланомы с эпидермальной сигнализацией [2]. Повышение экспрессии и активация EGFR и его лигандов (в частности, EGF) связано с проинвазивным фенотипом меланомы и резистентностью к терапии и опосредуется одним из механизмов поддержания клеток с фенотипом EGFR-положительной меланомы – регуляцией транскрипционным фактором NRF2 [9].
Следует обратить внимание на то, что выявленное более высокое содержание EGF (ключевой лиганд EGFR) в опухоли, ее перифокальной зоне и в ЛУ у мышей контрольной группы на момент окончания эксперимента сопровождалось метастатическим изменением подмышечных и паховых ЛУ, что говорит об агрессивности выросшей меланомы. Как показали результаты исследований A. Bracher и соавт., EGF играет важную роль в лимфангиогенезе и метастазировании меланомы в ЛУ [10], в работе K. Pietraszek-Gremplewicz и соавт. продемонстрировано прямое влияние уровня EGFR на инвазивный потенциал клеток меланомы [11], а B. Boone и соавт. была показана связь экспрессии EGFR в меланоме с ее метастатической инвазией в сторожевые лимфатические узлы [12].
Несмотря на то, что при меланоме рецепторные тирозинкиназы редко мутируют, известно, что около 6% меланом имеют активирующие мутации EGFR наряду с карциномой легкого и глиобластомой, а также примерно в половине случаев узловых меланом наблюдается экспрессия мембранного EGFR и одновременная экспрессия мембранных и ядерных рецепторов [13]. Кроме этого, стало известно, что активация сигнального пути EGFR связана с мутациями генов BRAF [14] и является одной из причин резистентности меланомы к анти-BRAF терапии препаратом вемурафениб [4], в связи с чем в настоящее время рассматриваются возможности использования таргетной терапии ингибиторами EGFR в схемах лечения больных меланомой [15]. Кроме этого, была продемонстрирована возможность одновременного воздействия на клетки меланомы комбинацией препаратов гефитиниба/лапатиниба (ингибиторы EGFR) и форетиниба (ингибитора MET – рецептора фактора роста гепатоцитов) с выраженным синергетическим эффектом подавления не только роста опухолевых клеток, но и их метастазирования [16].
Результаты экспериментальной терапии гефитинибом (группа сравнения) и веществом 6EPSVan-Na (основная группа) показали более медленное развитие опухоли на раннем сроке (3-е сутки после перевивки), на 30–60% меньший объем опухоли в конечной точке эксперимента (7-е сутки после окончания введения препаратов), чем у контрольных мышей, для вещества 6EPSVan-Na – снижение частоты синхронного поражения паховых и подмышечных ЛУ, обширные некротические изменения опухоли. Изменение течения меланомы у мышей с экспериментальной терапией сопровождалось в целом нормализацией содержания лиганда и растворимой формы его рецептора. Однако воздействие гефитинибом сопровождалось увеличением содержания EGF в ткани опухоли при его снижении в перифокальной зоне по сравнению с контролем, что может отражать локальное перераспределение фактора роста в связи с возможной адаптацией опухоли и повышение активности EGFR-зависимых сигнальных путей. Одновременно у мышей с введением 6EPSVan-Na, наоборот, отмечено снижение содержания EGF и в опухоли, и в перифокальной зоне по сравнению с контролем, при этом важно отметить, что в опухоли и в ЛУ оно оказалось статистически значимо меньше, чем в аналогичных тканях у мышей с гефитинибом. В совокупности такое изменение содержания EGF, возможно, способствовало более выраженному противометастатическому эффекту и некротическому поражению опухоли у мышей с введением 6EPSVan-Na по сравнению с влиянием гефитиниба.
Известно, что помимо полноразмерной трансмембранной формы рецептора EGFR в тканях (нормальных и опухолевых) и крови существуют его растворимые изоформы – sEGFR, содержащие только внеклеточный домен (сайт связывания лиганда EGF), образующиеся в результате альтернативного сплайсинга мРНК или протеолитического расщепления внеклеточного домена EGFR металлоферментами. Причем активация этих процессов – частое событие, регулирующее гомеостаз. Роль таких изоформ до сих пор однозначно не определена, зачастую противоречива [17]. Исследования показывают, что их уровень может отражать количество активированных полноразмерных рецепторов, чувствительных к ингибированию, то есть предположительно это маркер ответа на терапию ингибиторами EGFR. В статье A. Guillaudeau и соавт. приведен ряд работ, демонстрирующих возможность прогнозирования эффективности лечения в зависимости от уровня sEGFR не только в крови, но и в тканях опухолей при раке шейки матки, яичников, молочной железы, легких, колоректальном раке [18]. Кроме этого, предполагается, что sEGFR могут выполнять регуляторную роль, модулируя передачу сигнала полноразмерной формой EGFR посредством конкурентного связывания лигандов EGFR, образования неактивных гетеродимеров между разными изоформами [19]. Эти исследования подтверждают полученные результаты – более высокий, чем в коже интактных мышей, уровень sEGFR во всех изученных тканях мышей с ростом опухоли без воздействия (контрольная группа) и более низкий уровень sEGFR в тканях мышей после воздействия на опухоль гефитинибом или 6EPSVan-Na, чем в контрольной группе. Аналогичные изменения наблюдались и в изученных ЛУ. Такие результаты в целом демонстрируют однонаправленность и сопоставимость влияния гефитиниба и 6EPSVan-Na на уровень sEGFR, а в совокупности с более выраженным снижением содержания EGF в опухоли и лимфоузлах после воздействия 6EPSVan-Na свидетельствуют о перспективности дальнейшего изучения данного вещества.
Одним из возможных механизмов влияния sEGFR на модуляцию передачи сигнала трансмембранным рецептором EGFR называют конкурентное связывание лигандов EGFR, то есть sEGFR выполняет роль ловушки для EGF [19]. В ряде исследований in vitro было показано, что укороченные растворимые формы EGFR могут регулировать передачу сигналов этим рецептором в нормальных и опухолевых клетках, снижая пролиферацию [18]. Рассчитанное соотношение sEGFR/EGF дает упрощенное представление об изменении количества таких ловушек на одну молекулу лиганда. Анализ результатов говорит об их неоднозначности и нелинейности, при этом выделяются интересные изменения в перифокальной зоне опухоли – меньшее количество ловушек EGF в контрольной группе и после терапии гефитинибом и увеличение их числа после терапии 6EPSVan-Na (по сравнению с кожей интактных мышей), еще раз подчеркивая значимость метаболического состояния перифокальной зоны в патогенезе меланомы. По мнению авторов, интересные изменения отмечены и в ЛУ: рост величины sEGFR/EGF в подмышечных ЛУ после терапии обоими препаратами, особенно выраженный в паховых ЛУ после терапии 6EPSVan-Na, что может отражать увеличение количества ловушек EGF.
Влияние изменения уровня растворимой изоформы рецептора EGF в процессе лечения ингибиторами тирозинкиназ на развитие опухоли и в целом функциональная роль sEGFR еще далеки от понимания. Связывание sEGFR с лигандами EGFR и ограничение сигналов, усиливающих пролиферацию и способствующих опухолевой трансформации, а также снижение ее экспрессии при раке яичников, молочной железы, раке легких указывают на антионкогенную роль этой изоформы рецептора. С другой стороны, этому наблюдению противоречит связь повышенной экспрессии sEGFR при ряде других опухолей с более низкой общей выживаемостью, свидетельствуя, что регуляция экспрессии и роль изоформ sEGFR могут зависеть от типа опухоли. Необходимость понимания и объяснения этих различий требует дальнейших исследований [20]. В частности, глубокое изучение другого сигнального пути – фактора роста эндотелия сосудов (VEGF), являющегося важнейшим регулятором васкуло/ангио/лимфангиогенеза в норме и в патогенезе опухолевого роста, показало, что спектр растворимых изоформ рецепторов VEGFR представляет собой динамический регуляторный механизм роста сосудов в разных тканях и различных метаболических условиях, в зависимости от этого ингибируя или стимулируя ангиогенез, а в патологических условиях способствуя пролиферации опухоли и ее устойчивости к антиангиогенной терапии [21].
Заключение
Подводя итог, следует отметить, что разработанная и синтезированная в Пятигорском медико-фармацевтическом институте новая субстанция 6EPSVan-Na оказывает сопоставимое с гефитинибом противоопухолевое действие на меланому В16/F10, одновременно снижая частоту синхронного поражения паховых и подмышечных ЛУ. Вероятно, это может быть обусловлено более выраженным уменьшением содержания EGF в ткани опухоли и в метастатически пораженных подмышечных и паховых ЛУ, а также менее выраженным уменьшением содержания sEGFR в перифокальной зоне меланомы, что приводило к возрастанию величины sEGFR / EGF (предположительно отражающей увеличение количества ловушек EGF) после терапии 6EPSVan-Na.
Conflict of interest
Financing
Библиографическая ссылка
Франциянц Е.М., Сурикова Е.И., Бандовкина В.А., Кодониди И.П., Глушко А.А., Чиряпкин А.С., Шихлярова А.И., Алексеев Э.К., Шалашная Е.В., Нескубина И.В., Качесова П.С., Трепитаки Л.К., Петрова Ю.А., Черярина Н.Д., Солнцева А.А. ИЗМЕНЕНИЕ СОДЕРЖАНИЯ ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА И РАСТВОРИМОЙ ФОРМЫ ЕГО РЕЦЕПТОРА ПРИ АНТИМЕТАСТАТИЧЕСКОМ ДЕЙСТВИИ НОВОГО БЛОКАТОРА ВНУТРЕННЕГО ДОМЕНА EGFR– ПРОИЗВОДНОГО ПИРИМИДИН-4(1Н)-ОНА // Современные проблемы науки и образования. 2026. № 6. ;URL: https://science-education.ru/en/article/view?id=34640 (дата обращения: 14.07.2026).
DOI: https://doi.org/10.17513/spno.34640