


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
MORPHOFUNCTIONAL CHARACTERISTICS OF THE OVARY IN THE BACKGROUND OF DARK DEPRIVATION

Введение
По данным Росстата, за период с 2013 по 2023 год наблюдается значительное сокращение численности новорожденных детей в России: количество младенцев уменьшилось с 1357310 до 949742, что соответствует снижению показателя рождаемости на 46% [1]. Данная отрицательная статистика требует пристального внимания со стороны исследователей в области демографии, здравоохранения и социальной науки, поскольку отражает глубокие социальные перемены, обусловленные глобализацией: распространением единых стандартов образа жизни, включая увеличение доли ночных смен, круглосуточную доступность услуг и развлечений, рост уровня стресса на фоне высокой конкуренции и информационной перегрузки, и урбанизацией - ростом искусственного освещения в ночное время в городах. Световое загрязнение приводит к нарушению выработки мелатонина, являющегося ключевым регулятором биологических ритмов, циркадных ритмов на фоне хронического стресса, и гормональному дисбалансу [2; 3]. По данным Росстата, с 2020 по 2024 год наблюдается рост заболеваемости среди женщин, связанный с расстройством менструации на 37,3%, бесплодием на 7,8% [1]. Среди женщин репродуктивного возраста распространенность синдрома поликистозных яичников составляет от 8 до 21%, аменореи – от 5% до 13%, олигоменореи – от 8% до 22%, что связано с изменением уровня половых гормонов, нарушением менструального цикла, созреванием яйцеклеток, преждевременной недостаточностью яичников и ранним наступлением менопаузы [4-6].
Актуальность изучения морфофункциональных особенностей органов репродуктивной системы на фоне темновой депривации обусловлена необходимостью своевременного выявления, коррекции и профилактики преждевременной недостаточности яичников, что является важной задачей современной медицины.
Цель исследования. Выявить гистоморфометрические изменения в яичниках крыс при воздействии полной темновой депривации и коррекции экзогенным мелатонином.
Материал и методы исследования
Исследование проводилось в лаборатории токсикологии научного центра инновационных лекарственных средств с опытно-промышленным производством Волгоградского государственного медицинского университета (ВолгГМУ).
В качестве экспериментальных животных были выбраны 4-месячные половозрелые беспородные белые самки крыс, обладающие схожим с человеком морфофункциональным строением органов репродуктивной системы [7]. Общее количество животных составило 24 особи. Животные были получены из питомника Столбовая («Научный центр биомедицинских технологий», Россия). Все животные находились в одинаковых стандартных условиях: температура окружающей среды автоматически поддерживалась на уровне 25 °C. Крысы размещены в клетки по 6 особей в каждой с неограниченным доступом к гранулированному корму и питьевой воде. Эксперимент получил одобрение этического комитета ВолгГМУ (справка № 2022/164 от 25.11.2022).
После двухнедельного карантина, определения стадии эстрального цикла при помощи анализа влагалищных мазков перед началом эксперимента животные с одинаковыми фазами эстрального цикла были рандомно распределены на 2 группы: контрольную (n=6) и экспериментальную (ТД) (n=18). Животные контрольной группы содержались при стандартном автоматическом искусственном режиме освещения, предусматривающем чередование 12 ч света/12 ч темнота в течение 30 суток. Животные экспериментальной группы были размещены в помещении при автоматическом круглосуточном искусственном освещении (режим 24 ч света/0 часов темноты, 300 Люкс) на протяжении 30 суток [8]. На 31-е сутки животные экспериментальной группы 2 (ТД+14ЕО) и 3 (ТД+14ЕО+МТ) были возвращены в автоматическое стандартное искусственное освещение (режим 12 ч света/12 часов темноты). Животным группы ТД+14ЕО внутрижелудочно через зонд вводили 2% крахмальную слизь, ТД+14ЕО+МТ – мелатонин в дозе 0,3 мг/кг (Мелатонин-СЗ (3 мг), «Северная Звезда» НАО, Россия).
После завершении эксперимента на 31-е сутки животных контрольной и экспериментальной группы 1 (ТД), на 45-е сутки животных группы ТД+14ЕО и ТД+14ЕО+МТ взвешивали, затем под общей анестезией хлоралгидратом производили забор крови из брюшной аорты, извлекали яичники. Методом иммуноферментного анализа с использованием наборов ELISA Kit (Cloud-Clone Corp., США) на автоматическом микропланшетном фотометре Sunrise TS4TECAN (Tecan Austria GmbH, Австрия) проводили исследование сыворотки крови на содержание антимюллерова гормона.
Регистрацию масс яичников осуществляли на электронных весах ГОСМЕТР ВЛТЭ-1100С (ГОСМЕТР НПП, Россия). Максимальный предел взвешивания составлял 1100 г, цена деления 0,01 г. Класс точности II.
После предварительной фиксации яичников в 10%-ном растворе нейтрального забуференного формалина гистологические кассеты с материалом помещались в гистопроцессор Leica TP1020 (Leica Biosystems Nussloch GmbH, Германия) для проведения автоматической гистологической проводки с использованием программы № 6 – по спиртам возрастающей концентрации и ксилолом с последующей заливкой в гомогенизированную парафиновую среду «Гистомикс» («БиоВитрум», Россия). Серийные гистологические срезы толщиной 5 мкм изготавливали на ротационном микротоме Microm HM340E (Thermo Fisher Scientific, Германия), последовательно нарезались. Для исследования брался каждый 10-й срез [9], который монтировался на предметные стекла, обработанные поли-L-лизином (Menzel, Германия). Гистологические препараты окрашивались гематоксилином Майера и эозином по общепринятым методикам. Микроскопию и морфометрию гистологических препаратов проводили на микроскопе Leica DM1000 (Leica Microsystems GmbH, Германия) с применением программного обеспечения LAS 4.7.1. Выполняли по 10 снимков случайных полей зрения с каждого препарата для подсчета общего количества фолликулов. Для оценки риска преждевременного старения яичников на фоне стресса, вызванного темновой депривацией, осуществлялся подсчет растущих и зрелых фолликулов.
Статистическая обработка данных проводилась в программе GraphPad Prism 8.0.1. с использованием непараметрического дисперсионного анализа Краскела - Уоллиса с помощью теста Данна и параметрического One-way ANOVA с поправкой Тьюки. Результаты считали статистически значимыми при p < 0,05.
Результаты исследования и их обсуждение
Проведенное морфологическое исследование яичников на фоне 30-суточной темновой депривации в группе ТД выявило уменьшение массы яичников на 28,6% (p<0,05) по сравнению с контрольной группой. Площадь яичника уменьшилась на 24,9% (p<0,001) (рис.1).
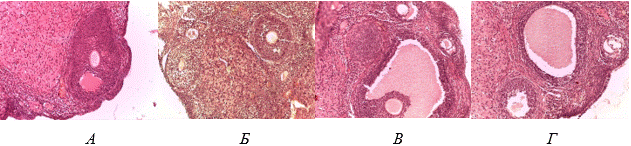
Рис. 1. Поперечный срез яичника крыс контрольной группы (А), после 30-суточной ТД (Б), после ТД+14 сут. ЕО (В), после ТД+14 сут. ЕО+МТ (Г). Увеличение числа атретических фолликулов, уменьшение количества функционально активных фолликулов по сравнению с контрольной (Б); увеличение числа растущих фолликулов по сравнению с группой ТД (В, Г). Окраска гематоксилином и эозином. Ув. х200
Выявлено нарушение нормальной структуры яичников. Это выражалось неравномерным распределением коркового и мозгового слоя. Отмечено увеличение толщины мозгового слоя яичника по сравнению с корковым слоем. Количество растущих и зрелых фолликулов снижено на 31,0% (p<0,001) в основном за счет третичных фолликулов на 42,8% (p<0,001) (рис. 2).
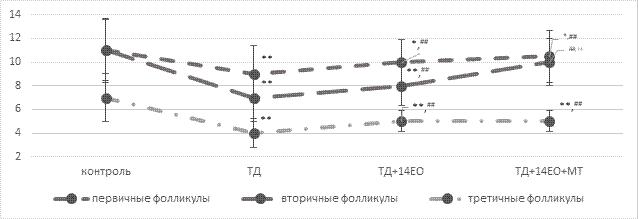
Рис. 2. Количественные данные фолликулов яичников под влиянием 30-дневной темновой депривации и коррекции в течение 14 суток естественным освещением и введением экзогенного мелатонина (Мe [25%;75%])
Примечание: составлено автором по результатам данного исследования;
* p<0,05 – по отношению к показателю контрольной группы,
**p<0,001 – по отношению к показателю контрольной группы,
##p<0,001 – по отношению к показателю группы ТД,
++p<0,001 – по отношению к показателю группы ТД+14ЕО (ранговый однофакторный анализ Краскела - Уоллиса, критерий Данна)
Через 2 недели после прекращения воздействия темновой депривации наблюдается положительная динамика в структуре яичников. Под влиянием эндогенного и введения экзогенного мелатонина отмечаются признаки регенерации ткани. Масса яичников в группах ТД+14ЕО и ТД+14ЕО+МТ увеличилась на 26,7% (p<0,05) и 33,3% (p<0,05) соответственно. Площадь яичника в группах ТД+14ЕО и ТД+14ЕО+МТ увеличилась на 15,6% (p<0,001) и 24,7% (p<0,001) соответственно.
Количественный анализ показал восстановление толщины коркового слоя яичников и уменьшение мозгового. В группах ТД+14ЕО отмечается статистически значимый рост (p<0,001) первичных, вторичных и третичных фолликулов по сравнению с группой ТД за счет активации фолликулогенеза из примордиальных фолликулов на 11,1%, 14,3% и 25% соответственно; в группе ТД+14ЕО+МТ – на 16,7%, 42,9% и 25,0% соответственно.
Особое внимание обращает на себя изменение уровня антимюллерова гормона при изменении свето-темнового режима. Проведенное исследование выявило статистически значимое изменение концентрации антимюллерова гормона в сыворотке крови крыс в виде его снижения на 30,9% (p<0,001) на фоне 30-суточной темновой депривации по сравнению с контрольной группой. Отмена темновой депривации на 14 суток и введение экзогенного мелатонина сопровождалось статистически значимым повышением уровня антимюллерова гормона в группах ТД+14ЕО и ТД+14ЕО+МТ на 41,4% (p<0,05) и на 33,7% (p<0,001) соответственно по сравнению с контрольной группой (рис. 3).
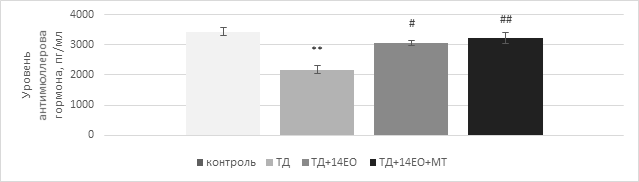
Рис. 3. Влияние 30-дневной темновой депривации на уровень антимюллерова гормона в сыворотке крови самок крыс (свето-темновой цикл 24/0 ч), M±m
Примечание: составлено автором по результатам данного исследования;
**p<0,001 - по отношению к показателю контрольной группы,
#p<0,05 - по отношению к показателю группы ТД,
##p<0,001 - по отношению к показателю группы ТД (One-way ANOVA с поправкой Тьюки)
Определение овариального резерва базируется на комплексном изучении количественных и качественных характеристик ооцитов. Среди ключевых методов косвенной оценки количества ооцитов выделяется определение уровня антимюллерова гормона и подсчет числа антральных фолликулов. Антимюллеров гормон начинает вырабатываться на ранних этапах развития ооцитов гранулёзными клетками преантральных и малых антральных фолликулов. Он играет важную роль в процессе отбора, обеспечивая контроль над скоростью активации примордиальных и преантральных фолликулов посредством аутокринных и паракринных механизмов. Благодаря этому антимюллеров гормон подавляет экспрессию ряда факторов роста, включая фактор роста фибробластов, фактора роста кератиноцитов и фактор стволовых клеток в фолликулах, замедляя темпы перехода фолликулов в фазу активного роста [10; 11].
Уменьшение концентрации антимюллерова гормона после 30-суточной темновой депривации приводит к увеличению скорости атрезии фолликулов, что подтверждается уменьшением числа растущих и зрелых фолликулов, и способствует снижению овариального резерва. Уровень антимюллерова гормона коррелирует с числом антральных фолликулов и выступает в качестве оценки фолликулярного пула. В связи с этим определение уровня содержания антимюллерова гормона в крови используется для прогнозирования преждевременной недостаточности яичников и является чувствительным маркером повреждения яичников [12]. Негативные последствия длительной темновой депривации сопровождаются снижением выработки мелатонина [13], что дополнительно усиливает процессы дегенерации фолликулов. Антимюллеров гормон снижает чувствительность первичных фолликулов к фолликулостимулирующему гормону [14]. Увеличение секреции фолликулостимулирующего гормона, наблюдаемое в результате нарушения нормального светового режима [15], подтверждает снижение функционального состояния яичников [16], несмотря на то, что данный гормон имеет меньшую диагностическую ценность по сравнению с антимюллеровым гормоном. Это обусловлено экспрессией антимюллерова гормона на фолликулах, у которых отсутствуют рецепторы к фолликулостимулирующему гормону. Уровень антимюллерова гормона относительно независим от уровня гонадотропинов [17].
Отмена 30-суточной темновой депривации, нормализация выработки эндогенного мелатонина или введение экзогенного мелатонина оказывают положительное влияние на репродуктивную систему. Наблюдается улучшение процесса созревания доминантного фолликула и развитие яйцеклетки за счет антиоксидантных свойств мелатонина [18], повышение уровня секреции антимюллерова гормона и эндогенного мелатонина [13], одновременно отмечается снижение секреции фолликулостимулирующего гормона [15; 19].
Согласно полученным нами данным по степени морфометрических изменений, мелатонин оказывает положительное влияние на регуляцию биоритмов и способствует физиологической выработке фолликулов, что также подтверждается данными Андреева Е. Н., Qi M. K., Yang C., Bisquert R. Мелатонин также участвует в регуляции биологических ритмов яичников, формировании фолликулов и задерживает старение яичников через путь мелатониновых рецепторов типа 1 (MT1)/AMP-активированной протеинкиназы, восстановление ДНК, поддержание теломер, белки сиртуины (SIRT1), регулирующие пролиферацию и апоптоз гранулезных клеток [20]. Мелатонин, регулируя путь PI3K/AKT, подавляет раннюю атрезию фолликулов яичников, тем самым замедляет истощение фолликулярного резерва [21]. Биохимический механизм влияния мелатонина реализуется благодаря активации сигнального пути Hippo, который регулирует баланс оксидативных реакций и антиоксидантных систем, поддерживая образование фолликулов, пролиферацию и дифференцировку гранулезных клеток, что необходимо для развития и функционирования яичников [20]. За счет этого достигается улучшение гормонального фона: нормализуется продукция гормонов базофильными клетками аденогипофиза, таких как лютеинизирующий и фолликулостимулирующий гормоны, которые контролируют процессы фолликулогенеза [19; 22], ослабляется окислительный стресс и усиливается антиоксидантная защита организма [23]. Эти механизмы обеспечивают созревание доминантного фолликула даже при изначально низком овариальном резерве и преждевременной недостаточности яичников [18; 19].
Заключение
Световой десинхроноз в виде темновой депривации оказывает негативное воздействие на морфофункциональное состояние яичников крыс. Продолжительная депривация естественного цикла освещения на протяжении 30 суток ускоряет дегенеративные процессы в тканях яичников, вызывая преждевременную недостаточность яичников. Уровень антимюллерова гормона совместно со снижением числа растущих фолликулов является маркером преждевременной недостаточности яичников. Темновая депривация проявляется снижением уровня антимюллерова гормона и уменьшением числа растущих фолликулов, негативно влияя на репродуктивное здоровье женского организма. Эндогенный и экзогенный мелатонин оказывает положительное воздействие на морфологическую картину яичников. Повышение секреции эндогенного мелатонина и введение экзогенного мелатонина оказывает стимулирующее влияние на фолликулогенез, обеспечивает созревание доминантного фолликула и опосредованно влияет на повышение уровня антимюллерова гормона.
Conflict of interest
Financing
Библиографическая ссылка
Кондакова Л.И. МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ЯИЧНИКОВ НА ФОНЕ ТЕМНОВОЙ ДЕПРИВАЦИИ // Современные проблемы науки и образования. 2026. № 5. ;URL: https://science-education.ru/en/article/view?id=34598 (дата обращения: 29.07.2026).
DOI: https://doi.org/10.17513/spno.34598