


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
THE GENES REGULATING MITOCHONDRIAL FUNCTIONING COPY NUMBER VARIATION FEATURES IN UTERINE CANCER PATIENTS WITH TYPE 2 DIABETES MELLITUS

Введение. Рак тела матки (РТМ) занимает лидирующие позиции в структуре онкогинекологической заболеваемости [1]. Одним из значимых коморбидных состояний при РТМ является сахарный диабет 2 типа (СД2), который выявляется у 15-30% пациенток [2]. Митохондриальная дисфункция играет ключевую роль в патогенезе как онкологических заболеваний, так и СД2 [3; 4]. Копийность генов (copy number variation, CNV), представляющая вариацию количества копий определенных генов в геноме, является важным молекулярным механизмом, влияющим на функционирование митохондрий. CNV возникают в результате делеций, дупликаций, инверсий и других структурных перестроек ДНК и могут иметь значительные функциональные последствия, включая изменения уровня экспрессии генов, нарушения в работе белков и развитие заболеваний [5].
Исследования показывают, что CNV широко распространены в геноме человека и ассоциированы с различными заболеваниями, включая нейродегенеративные, метаболические и онкологические патологии, в том числе рак эндометрия [6; 7].
Изменения копийности генов, регулирующих функционирование митохондрий, могут иметь важное значение в молекулярных механизмах развития и прогрессирования РТМ [8], влияя на апоптоз, энергетический метаболизм и устойчивость к терапии [9]. При этом наличие СД2 может усугублять эти нарушения, способствуя прогрессированию опухоли и снижению эффективности лечения [10; 11].
Целью данного исследования явился анализ относительной копийности генов, регулирующих функционирование митохондрий, у больных РТМ с СД2 и без него.
Материалы и методы исследования
На биоинформационном этапе исследования был выполнен анализ открытой базы данных TCGA (The Cancer Genome Atlas, https://www.cancer.gov/ccg/research/genome-sequencing/tcga) для выявления при раке тела матки аномальной копийности генов, ассоциированных с митохондриальной дисфункцией и сахарным диабетом 2 типа. Данные о числе копий генов были получены из Genomic Data Commons Data Portal (https://portal.gdc.cancer.gov/) с помощью TCGABiolinks (R v.4.0.0, Rstudio). Алгоритм GISTIC был использован для обнаружения областей генома, размер которых значимо изменялся в ряде опухолевых образцов [12].
В исследование была включена 71 пациентка с морфологически верифицированным диагнозом «аденокарцинома тела матки». Больные получали лечение (хирургическое, а также адъювантная терапия: лучевая или комбинации химио- и лучевой терапии) в период с 2023 по 2025 год. Средний возраст пациенток составил 58,3±8,7 года. В зависимости от степени дифференцировки пациентки были разделены на три группы: G1 (n=20), G2 (n=35) и G3 (n=16) [13]. В каждой группе выделяли подгруппы с сопутствующим СД2 и без него. Диагноз СД2 устанавливался на основании критериев ВОЗ (2019). В группе G1 было 6 пациенток с СД2 и 14 без СД2), в группе G2 - 14 пациенток с СД2 и 21 без СД2, в группе G3 - 6 пациенток с СД2 и 10 без СД2.
Относительная копийность генов определялась методом количественной полимеразной цепной реакции (qPCR) в реальном времени в образцах ДНК (выделяли методом фенол-хлороформной экстракции) опухолевой и условно нормальной ткани, полученных при биопсии или во время операции. Анализировались гены, ассоциированные с митохондриальной функцией: MT-CO1, mtTFB, mtSSB, ATAD3, TFAM, POLG, POLRMT, TOP1MT, MGME1, TEFM, MT-RNR2, GCAT и NRF1. В качестве референсных использовались гены домашнего хозяйства (GAPDH, ACTB). Реакцию амплификации проводили на термоциклере CFX96 (Bio-Rad, США) в 20 мкл ПЦР-смеси (ДНК (не менее 1 нг), 0,2 мМ dNTP, 0,4 мкМ праймеры, 2,5 мМ MgCl2, 1X ПЦР-буфер с интеркалирующим красителем EvaGreen и 0,1 е.а./мкл SynTaq ДНК-полимеразы), по следующей программе: 95 °C 4 минуты, 40 циклов: 95 °C 10 секунд, 58 °C 30 секунд (чтение оптического сигнала по каналу FAM) и 72 °C 15 секунд. Относительную копийность (rC) рассчитывали по формуле: rC = E-ΔΔCt [14].
Статистическая обработка данных проводилась с использованием программы R v.4.3.1. Нормальность распределения показателей оценивали с помощью критерия Колмогорова - Смирнова. Для сравнения количественных показателей между группами применяли критерий Манна - Уитни c поправкой Бонферрони на множественное сравнение [14]. Различия считались статистически значимыми при p <0,05. Для описания изменения копийности генов в зависимости от наличия СД2 строились линейные регрессионные модели вида Y=β0+β1⋅X, где Y - относительная копийность гена, X - бинарный показатель наличия СД2 (0 - отсутствие, 1 - наличие), β0 - медиана копийности в группе без СД2, β1 - медиана копийности при наличии СД2.
Результаты исследования и их обсуждение
Проведенный биоинформационный анализ открытых баз данных позволил выявить ряд генетических локусов, изменение копийности которых ассоциировано с митохондриальной дисфункцией и сахарным диабетом 2 типа при раке тела матки - MT-CO1, mtTFB, mtSSB, ATAD3, TFAM, POLG, POLRMT, TOP1MT, MGME1, TEFM, MT-RNR2, GCAT, NRF1. Информация о функциях некоторых локусов представлена в таблице 1.
Таблица 1
Функции генов и клиническая значимость при патологиях матки
|
Ген |
Локализация |
Основная функция |
Роль в онкогенезе РТМ |
|
MT-CO1 |
Митохондрии |
Кодирует субъединицу цитохром-c-оксидазы (каталитическая субъединица комплекса IV дыхательной цепи) |
Снижение → нарушение OXPHOS → активация HIF-1α |
|
mtSSB |
Митохондрии |
Связывание одноцепочечной мтДНК. Стабилизирует одноцепочечную мтДНК при репликации |
Снижение → дестабилизация мтДНК |
|
MT-RNR2 |
Митохондрии |
Кодирует 16S рРНК |
Снижение → подавление синтеза митохондриальных белков |
|
MGME1 |
Ядро/ митохондрии |
Эндонуклеаза репарации мтДНК, разрешает структуры flap мтДНК при репарации |
Противоречивая: гиперэкспрессия в G1/G2 → компенсация повреждений; гипоэкспрессия в G3 → коллапс мтДНК |
|
ATAD3 |
Ядро |
Регуляция стабильности мтДНК, ядерно-митохондриальный «якорь» для транспорта холестерина |
Поддержание базовых функций митохондрий |
|
TFAM |
Ядро |
Фактор транскрипции мтДНК |
Сохранение биогенеза мтДНК |
|
POLG |
Ядро |
ДНК-полимераза γ |
Репликация мтДНК |
Примечание: составлено авторами на основе источников [3; 4; 9], а также результатов биоинформационного анализа.
Показатель копийности данных генетических локусов был валидирован методом ПЦР-РВ на образцах ДНК из тканей 71 пациентки с аденокарциномой тела матки.
У пациенток с аденокарциномой G1 без СД2 наблюдалось статистически значимое (p<0.005) снижение копийности мтДНК-связанных генов (MT-CO1=0.6, mtTFB=0.5, mtSSB=0.6, MT-RNR2=0.5), снижение NRF1(0.5) и повышение MGME1 (2.5) относительно условно-нормальной ткани. У пациенток с аденокарциномой G1 и СД2 наблюдалось статистически значимое (p<0.05) более выраженное снижение копийности мтДНК-связанных генов (MT-CO1=0.5, mtTFB=0.5, mtSSB=0.4, MT-RNR2=0.3) и ядерных генов NRF1(0.5) и TFAM (1.6) относительно условно-нормальной ткани (рис. 1). При этом следует отметить, что в группах пациенток с сахарным диабетом и без статистически значимо (p<0.05) отличалась копийность 2 генов: копийность TFAM и NRF1 была в 2,5 раза меньше у пациенток с сахарным диабетом (рис. 1).
У пациенток с аденокарциномой G2 без СД2 наблюдалась схожая картина с G1: статистически значимое (p<0.005) снижение копийности мтДНК-генов (MT-CO1=0.7, mtTFB=0.7, MT-RNR2=0.6), кроме mtSSB (1.4 - ближе к норме) и NRF1(0.5). Кроме этого, наблюдалось статистически значимое (p<0.005) повышение копийности MGME1 (2.9). У пациенток с аденокарциномой G2 и СД2 наблюдалось статистически значимое (p<0.005) более выраженное снижение копийности мтДНК-генов (MT-CO1=0.4, mtTFB=0.3, mtSSB = 0,6, MT-RNR2=0.3). Также наблюдалось статистически значимое (p<0.005) снижение копийности ядерных генов - TFAM и TOP1MT в 2 раза и NRF1 в 5 раз относительно условно-нормальной ткани (рис. 2).
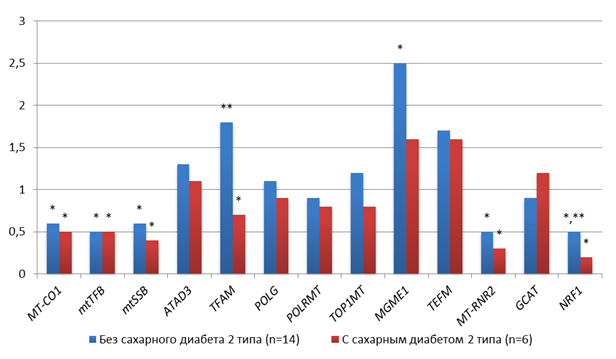
Рис. 1. Особенности копийности генов в тканях пациенток с аденокарциномой G1 с СД2 и без; * - статистически значимое отличие от условно-здоровой ткани, ** - статистически значимое отличие от пациентов с сахарным диабетом
Источник: составлено авторами по результатам данного исследования.
При этом следует отметить, что в группах пациенток с сахарным диабетом и без статистически значимо (p<0.05) отличалась копийность 7 генов: у пациенток с сахарным диабетом копийность TFAM была в 3,2 раза меньше, MT-CO1 в 1,8 раза меньше, mtTFB в 2,3 раза меньше, mtSSB в 2,3 раза меньше, TOP1MT в 1,8 раза меньше, MT-RNR2 в 2 раза меньше и NRF1 в 2,5 раза меньше, чем у пациенток без сахарного диабета (рис. 2).
У пациенток с аденокарциномой G3 без СД2 наблюдалось снижение копийности mtSSB и MT-RNR2 в 5 и 2.5 раза соответственно относительно условно-нормальной ткани (p<0.005), а также снижение копийности MGME1 в 1.4 раза (p<0.005), что противоположно результатам при G1/G2. У пациенток с аденокарциномой G3 и СД2 наблюдалось снижение копийности MT-CO1, mtTFB, mtSSB, MT-RNR2, NRF1, TFAM, TOP1MT и MGME1 в 2.5 раза, 3.3 раза, 5.0 раз, 3.3 раза, 1.7 раза, 1.7 раза и 5.0 раз соответственно относительно условно-нормальной ткани (p<0.005) (рис. 3). При этом следует отметить, что в группах пациенток с сахарным диабетом и без статистически значимо (p<0.05) также отличалась копийность 7 генов: у пациенток с сахарным диабетом копийность TFAM была в 2,5 раза меньше, MT-CO1 в 2,3 раза меньше, mtTFB в 4,0 раза меньше, TOP1MT в 2,2 раза меньше, MT-RNR2 в 2 раза меньше, NRF1 в 2,7 раза меньше и MGME1 в 3,5 раза меньше, чем у пациенток без сахарного диабета (рис. 3).
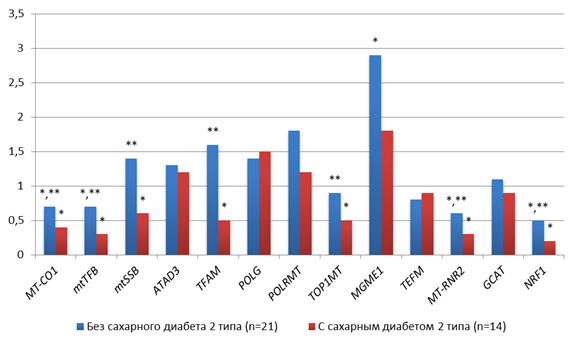
Рис. 2. Особенности копийности генов в тканях пациенток с аденокарциномой G2 с СД2 и без; * - статистически значимое отличие от условно-здоровой ткани, ** - статистически значимое отличие от пациентов с сахарным диабетом
Источник: составлено авторами по результатам данного исследования.
При всех трех степенях дифференцировки РТМ наблюдалось статистически значимое снижение копийности ряда митохондриальных и ядерных (ассоциированных с регуляцией митохондрий) генов (табл. 2). Следует отметить, что увеличение числа негативных аномалий копийности генов у пациенток с РТМ было выражено значительно сильнее при наличии СД2 типа и снижении степени дифференцировки клеток. Наиболее выраженные изменения отмечались для генов: MT-CO1: снижение на 16,7-55,6%, TFAM: снижение на 60,0-68,8%, NRF1: снижение на 60,0-62,5%.
На основе регрессионного анализа были построены уравнения, описывающие влияние СД2 на копийность генов, например: для гена TFAM в группе G1: Y = 1,8 - 1,1·X; для гена MT-CO1 в группе G2: Y = 0,7 - 0,3·X; для гена NRF1 в группе G3: Y = 0,8 - 0,5·X. Эти уравнения показывают, что наличие СД2 ассоциировано со снижением копийности данных генов на 0,3-1,1 условной единицы. Полученные результаты демонстрируют значительное влияние СД2 на копийность генов, отвечающих за функционирование митохондрий у пациенток с РТМ. Это согласуется с известными данными о том, что CNV могут влиять на экспрессию генов и способствовать развитию заболеваний [5]. Наиболее выраженные изменения выявлены при G2 и G3, что может указывать на Grade-специфичное влияние СД2 на митохондриальную дисфункцию при онкологических заболеваниях и может свидетельствовать о различной чувствительности пациентов к метаболическим нарушениям, ассоциированным с СД2 [15; 16].
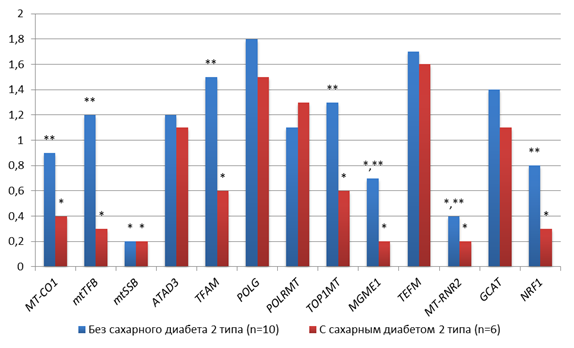
Рис. 3. Особенности копийности генов в тканях пациенток с аденокарциномой G3 с СД2 и без; * - статистически значимое отличие от условно-здоровой ткани, ** - статистически значимое отличие от пациентов с сахарным диабетом
Источник: составлено авторами по результатам данного исследования.
Таблица 2
Изменение копийности генов у пациенток с РТМ в зависимости от наличия СД2
|
Ген |
Группа |
Без СД2 |
С СД2 |
Изменение (%) |
p-value |
|
MT-CO1 |
G1 |
0,6* |
0,5* |
-16,7 |
0,045 |
|
MT-CO1 |
G2 |
0,7* |
0,4* |
-42,9 |
0,003 |
|
MT-CO1 |
G3 |
0,9* |
0,4* |
-55,6 |
0,001 |
|
TFAM |
G1 |
1,8* |
0,7* |
-61,1 |
0,002 |
|
TFAM |
G2 |
1,6* |
0,5* |
-68,8 |
0,001 |
|
TFAM |
G3 |
1,5* |
0,6* |
-60,0 |
0,004 |
|
NRF1 |
G1 |
0,5*,** |
0,2* |
-60,0 |
0,002 |
|
NRF1 |
G2 |
0,5*,** |
0,2* |
-60,0 |
0,001 |
|
NRF1 |
G3 |
0,8** |
0,3* |
-62,5 |
0,002 |
Источник: составлено авторами на основе полученных данных в ходе исследования.
Снижение копийности гена TFAM, играющего ключевую роль в репликации и транскрипции митохондриальной ДНК, может объяснять нарушение энергетического метаболизма в опухолевых клетках при сопутствующем СД2 [4; 9]. Аналогичные изменения копийности MT-CO1, кодирующего ключевой фермент дыхательной цепи, могут приводить к нарушению окислительного фосфорилирования и усилению гликолиза (эффект Варбурга) [3; 17]. Особого внимания заслуживает снижение копийности гена NRF1, который регулирует экспрессию ядерных генов, кодирующих митохондриальные белки. Это может приводить к дисбалансу в синтезе субъединиц электронтранспортной цепи и дальнейшему ухудшению функции митохондрий [9]. Метод qPCR, использованный в исследовании, является надежным инструментом для определения относительной копийности генов, однако он имеет ограничения, связанные с возможностью изучать небольшое количество локусов одновременно. В дальнейшем для более масштабного анализа CNV могут быть применены методы высокопроизводительного секвенирования [18].
Выявленные закономерности согласуются с данными литературы о том, что СД2 ассоциирован с более агрессивным течением РТМ и худшим ответом на терапию [11; 19]. Полученные данные подчеркивают важность учета коморбидного фона при выборе тактики лечения пациенток с РТМ [1].
Заключение
Наличие СД2 у пациенток с РТМ ассоциировано со статистически значимым снижением копийности генов, регулирующих функционирование митохондрий (MT-CO1, TFAM, NRF1). Наиболее выраженные изменения наблюдаются при степени дифференцировки опухолевых клеток G2 и G3. Построенные регрессионные модели количественно описывают ассоциацию СД2 с аномальной копийностью ядерных и митохондриальных генов и могут быть использованы для прогнозирования молекулярных последствий коморбидной патологии. Полученные данные обосновывают необходимость разработки персонализированных подходов к терапии РТМ с учетом метаболического статуса пациенток.
Перспективным направлением дальнейших исследований является изучение влияния выявленных изменений копийности генов на эффективность различных вариантов терапии РТМ, а также оценка возможности коррекции митохондриальной дисфункции как потенциального терапевтического подхода. Кроме того, перспективным является анализ CNV в циркулирующей ДНК плазмы крови как малоинвазивного биомаркера.
Conflict of interest
Библиографическая ссылка
Кутилин Д.С., Франциянц Е.М., Нескубина И.В., Шихлярова А.И., Каплиева И.В., Бандовкина В.А., Качесова П.С., Шейко Е.А., Гусарев А.Д., Енгибарян В.А., Удаленкова И.А., Озеркова Е.А. ОСОБЕННОСТИ КОПИЙНОСТИ ГЕНОВ, РЕГУЛИРУЮЩИХ ФУНКЦИОНИРОВАНИЕ МИТОХОНДРИЙ, У БОЛЬНЫХ РАКОМ ТЕЛА МАТКИ С САХАРНЫМ ДИАБЕТОМ 2 ТИПА // Современные проблемы науки и образования. 2025. № 5. ;URL: https://science-education.ru/en/article/view?id=34316 (дата обращения: 25.07.2026).
DOI: https://doi.org/10.17513/spno.34316