


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
RESEARCH ON C22orf20 GENE IN PATIENTS WITH TYPE 2 DIABETES MELLITUS AND ACCOMPANYING HEPATIC PATHOLOGY IN YAKUTIA

Сахарный диабет 2-го типа (СД 2) является одним из социально наиболее значимых патологий в Республике Саха (Якутия). Снижение чувствительности тканей к инсулину (инсулинорезистентность), которое является основой заболевания СД 2-го типа, приводит к нарушениям углеводного и липидного обмена всего организма и в первую очередь печени, что в свою очередь зачастую ведет к развитию тяжелых заболеваний печени [1]. Многочисленные исследования показывают частое сочетание СД 2-го типа и неалкогольной жировой болезни печени (НАЖБП), которая характеризуется накоплением липидов как в самих гепатоцитах, так и в межклеточном пространстве [2]. Распространенность НАЖБП среди пациентов СД 2-го типа составляет 60–80%, а частота развития НАСГ – 12–40% [3, 4]. Патогенетические механизмы НАЖБП и СД 2-го типа тесно взаимосвязаны. Сочетание данных заболеваний отягощает течение друг друга, значительно повышая вероятность развития у пациента фиброза печени. Является ли НАЖБП следствием или причиной метаболической дисфункции, в настоящее время неизвестно. Заболевания печени на ранней стадии протекают с невыраженной симптоматикой или бессимптомно, поэтому необходима своевременная диагностика и профилактика заболеваний печени, в особенности у пациентов с СД 2-го типа. Изучение молекулярно-генетического механизма патогенеза НАЖБП будет способствовать поиску новых биомаркеров заболевания [4].
Одним из кандидатныx генов, вовлеченных в патогенез НАЖБП, является пататинподобный фосфолипазный домен-содержащий белок 3 (C22orf20, PNPLA3, ADPN,), также известный как адипонутрин. Адипонутрин является триацилглицерин липазой, которая опосредует гидролиз триацилглицерина в адипоцитах и может играть определенную роль в энергетическом обмене. Полиморфизм этого гена является предиктором прогрессирующего течения НАЖБП и основным фактором риска трансформации НАЖБП в цирроз. Исследования показывают, что полиморфизм I148M может представлять общий модификатор фиброгенеза при заболеваниях печени [5].
Целью нашего исследования явилось изучение распределения частот аллелей среди русского и якутского населения Республики Саха (Якутия), а также поиск ассоциаций полиморфного варианта гена C22orf20 (rs738409 C>G) с заболеваниями печени у пациентов, страдающих СД 2-го типа.
Материал и методы исследования
Исследования полиморфизма I148M гена C22orf20 проводились в лаборатории наследственной патологии отдела молекулярной генетики Якутского научного центра комплексных медицинских проблем (ЯНЦ КМП). Для исследования использованы образцы ДНК из коллекции биоматериала ЯНЦ КМП. Выборка исследованных лиц состояла из 178 пациентов эндокринологического отделения Республиканской больницы № 2 Государственного бюджетного учреждения «Центр экстренной медицинской помощи» с диагнозом «Сахарный диабет 2-го типа» и сопутствующими заболеваниями печени неинфекционного генеза. В состав выборки вошли 27 пациентов русской и 151 пациент якутской национальностей. Критериями включения в исследование были: отсутствие поражения печени хроническими вирусными гепатитами; у всех исследуемых были исключены: аутоиммунный гепатит, первичный билиарный холангит, первичный склерозирующий холангит, наследственный гемохроматоз, болезнь Вильсона–Коновалова; отсутствие злоупотребления алкоголем (> 30 г/л).
Группой сравнения служила выборка из 135 здоровых добровольцев русской национальности и 246 здоровых добровольцев якутской национальности. В исследование были включены участники, заполнившие анкету, одобренную Локальным комитетом по биомедицинской этике при ЯНЦ КМП и добровольно подписавшие информированное согласие на проведение генетического исследования. Для участников, не достигших совершеннолетия, заполнение анкет и подписание информированного согласия осуществляли родители. Этническая принадлежность учитывалась до третьего поколения.
Выделение геномной ДНК проводилось стандартным методом фенольно-хлороформной экстракции из замороженной цельной крови. Генотипирование однонуклеотидного полиморфизма (SNP) I148M (rs738409) образцов ДНК осуществляли путем анализа продуктов полимеразной цепной реакции (ПЦР) – амплификации специфических участков генома с последующим анализом полиморфизма длин рестрикционных фрагментов (ПДРФ).
Условия проведения амплификации области гена, содержащего полиморфный вариант, с указанием последовательности олигонуклеотидных праймеров, используемой рестриктазы и длин рестрикционных фрагментов, представлены в таблице 1.
Таблица 1
Условия проведения амплификации
|
Праймер |
Условия проведения ПЦР |
Длина амплификата |
Рестриктаза |
Длина фрагментов рестрикции |
|
F: 5’-TGGGCCTGAAGTCCGAGGGT-3’ |
1. 95°С – 5 мин 2.(94°С – 30 сек; 66°С – 30 сек; 72°С – 40 сек)*37 3. 72°С – 5 мин |
333 п.н. |
BstF5 I |
CC – 200, 133 CG – 333, 200, 133 GG – 333 |
|
R: 5’- CCGACACCAGTGCCCTGCAG-3’ |
Определение генотипов проводилось анализом размеров образующихся фрагментов (рестриктов) путем гель-электрофореза на 4%-ном агарозном геле с бромистым этидием в стандартном трис-ацетатном буфере при 120 В в течение 1 часа. Визуализацию рестрикционных продуктов проводили c использованием гель-документирующей системы в UV-лучах (Vilber Lourmat, Франция) (рис).
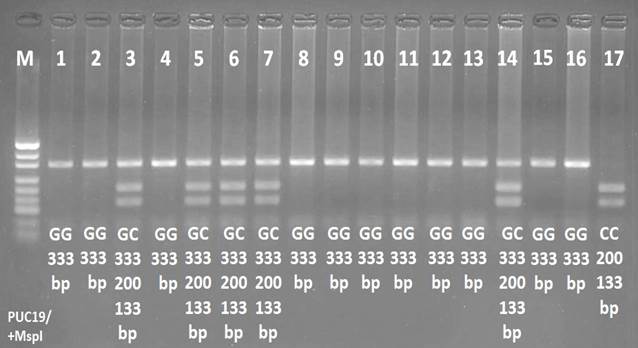
Электрофореграмма результатов ПЦР-ПДРФ анализа М – маркер PUC19/+Msp I. 1, 2, 4, 8, 9, 10, 11, 12, 13, 15, 16 – генотип GG, 3, 5, 6, 7, 14 – генотип GC, 17 – генотип CC. bp – пар оснований
Обработку данных исследования проводили статистическими программами «Office Microsoft Excel 2010» и «Statistica 8.0». Частоту аллелей и генотипов полиморфизма rs738409 гена C22orf20 проверяли на соответствие равновесию Харди–Вайнберга (РХВ) с помощью точного теста Фишера. Частоту аллелей и генотипов между выборками русских и якутов, как больных, так и здоровых, сравнивали с использованием критерия χ2 с поправкой Йетса на непрерывность. Результаты считались значимыми при значении р меньше 0,05 (p<0,05).
Результаты и обсуждение
Анализ распределения частот аллелей и генотипов полиморфного варианта гена C22orf20 (rs738409) в изученных выборках показал различие между якутами и русскими как у здоровых добровольцев, так и среди страдающих заболеваниями печени (табл. 2).
Таблица 2
Частота встречаемости аллелей и генотипов полиморфизма rs738409 гена C22orf20 у русских и якутов
|
Группы |
Кол-во |
Распр |
Генотип, % |
Аллель |
х2 |
р |
|||
|
CC |
CG |
GG |
С |
G |
|||||
|
Русские, популяционная выборка |
135 |
Н |
40,0 |
26,7 |
33,3 |
0,533 |
0,467 |
29,101 |
0,000 |
|
О |
28,4 |
49,8 |
21,8 |
||||||
|
Якуты, популяционная выборка |
246 |
Н |
13,8 |
26,4 |
59,8 |
0,270 |
0,730 |
26,825 |
0,000 |
|
О |
7,3 |
39,4 |
53,2 |
||||||
|
Больные якутской национальности |
151 |
Н |
9,9 |
29,8 |
60,3 |
0,248 |
0,752 |
6,147 |
0,013 |
|
О |
6,2 |
37,3 |
56,5 |
||||||
|
Больные русской национальности |
27 |
Н |
33,3 |
25,9 |
40,7 |
0,463 |
0,537 |
6,185 |
0,013 |
|
О |
21,4 |
49,7 |
28,8 |
||||||
Примечание: Н – наблюдаемое распределение, О – ожидаемое распределение
Выборка здоровых пациентов русской национальности характеризовалась наибольшим количеством носителей гомозиготного генотипа СС, тогда как в выборках пациентов с СД 2-го типа преобладали носители гомозиготного генотипа GG. Среди пациентов с СД 2-го типа и здоровой выборки якутской национальности наибольшее количество было носителей генотипа GG.
При сравнении частоты аллелей и генотипов у здоровых добровольцев якутской и русской национальностей имеется достоверное различие как между частотами генотипов, так и между аллелями (табл. 3).
Таблица 3
Сравнение частот генотипов и аллелей среди здоровых выборок русских и якутов
|
Генотипы, аллели |
Русские (n = 135) |
Якуты (n = 246) |
Х2 |
OR (95% CI) для аллелей |
Значимость, p |
|
С/С |
54 (40%) |
34 (13,8%) |
37,942 |
3,085 (2,260–4,210) |
0,000* |
|
С/G |
36 (26,7%) |
65 (26,4%) |
|||
|
G/G |
45 (33,3%) |
147 (59,8%) |
|||
|
С |
0,533 |
0,270 |
50,989 |
0,000** |
|
|
G |
0,467 |
0,730 |
Примечание. Достигнутый уровень значимости при сравнении распределения генотипов (*) и частоты аллелей (**) в группах сравнения 1 и 2. n – численность выборок, Х2 с поправкой Йетса
В выборке здоровых добровольцев русской национальности преобладали встречаемость частоты аллеля С и гомозиготный генотип СС, тогда как в выборке здоровых добровольцев якутской национальности преобладали частота аллеля G и гомозиготный генотип GG. Расчет отношения шансов показал, что частота встречаемости аллеля G в группе якутов достоверно выше (OR – 3,085; 95% CI: 2,260 – 4,210; р<0,001), чем у русских.
При анализе распределения частоты генотипов и аллелей среди здоровой выборки русских и выборки пациентов с заболеваниями печени достоверных различий не обнаружено (табл. 4).
Таблица 4
Сравнение частот генотипов и аллелей среди больных и здоровых русских
|
Генотипы, аллели |
Больные (n = 27) |
Контрольная выборка (n = 135) |
Х2 |
OR (95% CI) для аллелей |
Значимость, p |
|
С/С |
9 (33,3%) |
54 (40%) |
0,619 |
1,326 (0,738–2,382) |
0,734* |
|
С/G |
7 (25,9%) |
36 (26,7%) |
|||
|
G/G |
11 (40,7%) |
45 (33,3%) |
|||
|
С |
0,463 |
0,533 |
0,633 |
0,426** |
|
|
G |
0,537 |
0,467 |
Примечание. Достигнутый уровень значимости при сравнении распределения генотипов (*) и частоты аллелей (**) в группах сравнения 1 и 2. n – численность выборок, Х2 с поправкой Йетса.
В выборке пациентов с СД 2-го типа с сопутствующими печеночными патологиями русской национальности преобладал гомозиготный генотип GG, что, вероятно, связано с малым количеством выборки (n=27). Среди выборки здоровых добровольцев русской национальности преобладал гомозиготный генотип СС.
При анализе распределения частоты генотипов и аллелей среди здоровой выборки якутов и выборки пациентов с СД 2-го типа с сопутствующими заболеваниями печени достоверных различий не обнаружено (табл. 5).
Таблица 5
Сравнение частот генотипов и аллелей среди больных и здоровых якутов
|
Генотипы, аллели |
Больные (n = 151) |
Контрольная выборка (n = 246) |
Х2 |
OR (95% CI) для аллелей |
Значимость, p |
|
С/С |
15 (9,9) |
34 (13,8) |
1,535 |
1,121 (0,808–1,557) |
0,464* |
|
С/G |
45 (29,8) |
65 (26,4) |
|||
|
G/G |
91 (60,3) |
147 (59,8) |
|||
|
С |
0,248 |
0,270 |
0,361 |
0,548** |
|
|
G |
0,752 |
0,730 |
Примечание. Достигнутый уровень значимости при сравнении распределения генотипов (*) и частоты аллелей (**) в группах сравнения 1 и 2. n – численность выборок, Х2 с поправкой Йетса
Из данных открытых источников проекта «1000 Genomes» [6] следует, что с высокой частотой аллель G встречается в популяциях Центральной и Южной Америк (перуанцы – 71,8%, мексиканцы – 55,5%, колумбийцы – 41%). Обращает внимание тот факт, что, чем выше частота аллеля G, тем выше процент коренного населения в популяции. Например, в популяции Пуэрто-Рико, где коренное индейское население составляет 11 %, частота аллеля G составляет 31,7%. У европейцев частота аллеля G в среднем 22,6%. Среди азиатов высока частота аллеля G у японцев (42,3%). Обладатели же самой низкой частоты аллеля G – африканцы (8,6–17,2%).
Заключение
В результате исследования установлено достоверное различие между частотами генотипов и аллелей гена C22orf20 (rs738409) у русских и якутов как среди здоровых добровольцев, так и у больных СД 2-го типа с заболеваниями печени. Частота встречаемости аллеля G в группе здоровых якутов была достоверно выше (OR – 3,085; 95% CI: 2,260–4,210; р<0,001), чем в группе здоровых русских. При анализе распределения частот генотипов и аллелей среди здоровой выборки и выборки пациентов с заболеваниями печени как в русской, так и в якутской популяциях достоверных различий не обнаружено.
Исследование было проведено в рамках НИР Изучение генетической структуры и груза наследственной патологии популяций Республики Саха (Якутия) с использованием УНУ «Геном Якутии» (рег.№ USU_507512).
Библиографическая ссылка
Куртанов Х.А., Павлова Н.И., Дьяконова А.Т., Соловьева Н.А., Соловьева Ю.А., Сыдыкова Л.А., Филиппова Н.П., Александрова Т.Н. ИССЛЕДОВАНИЕ ГЕНА C22orf20 У БОЛЬНЫХ САХАРНЫМ ДИАБЕТОМ 2-ГО ТИПА С СОПУТСТВУЮЩЕЙ ПЕЧЕНОЧНОЙ ПАТОЛОГИЕЙ В ЯКУТИИ // Современные проблемы науки и образования. 2019. № 3. ;URL: https://science-education.ru/en/article/view?id=28768 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/spno.28768