


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
EFFECT OF HYPOTHERMIA ON ERYTHROCYTE NA, K-ATPASE ACTIVITY AND THE BALANCE OF SODIUM AND POTASSIUM IONS IN RAT’S PLASMA AND RED BLOOD CELLS UNDER DALARGIN INJECTION

Искусственное снижение температуры тела как метод снижения кислородных запросов организма успешно применяется для повышения устойчивости органов и тканей к различным видам стресса, защиты организма от травматических повреждений, восстановления функции органов и тканей после ишемии-реперфузии [3]. Наряду с повышением устойчивости организма к воздействию многих неблагоприятных факторов, гипотермия вызывает ряд нежелательных изменений. При снижении температуры тела происходит нарушение микроциркуляции, обусловленное вазоспазмом и централизацией кровотока, а также повышением вязкости крови [8]. Негативными эффектами гипотермии являются также смещение прооксидантно-антиоксидантного баланса в сторону избыточной генерации свободных радикалов и возникновение дефицита антиоксидантов [12]. Это в свою очередь, может быть причиной изменения физико-химического состояния эритроцитарных мембран [8] и нарушения работы мембраносвязанных ферментов.
Na, K-АТФаза является интегральным ферментом мембран эритроцитов, участвующим в регуляции ионного гомеостаза красных клеток. Ранее на изолированных мембранах эритроцитов крыс нами было установлено, что глубокая гипотермия (20 °С) снижает активность Na, K-АТФазы [6]. При гипотермии увеличивалось количество связанного с мембранами эритроцитов гемоглобина. Снижение активности Na, K-АТФазы при гипотермии отрицательно коррелировало с количеством связанного с мембранами гемоглобина. В процессе получения белых теней эритроцитов мембрана теряет ряд белков, особенно цитоскелета [5], что может отразиться на активности Na, K-АТФазы. В связи с этим представляет интерес исследование активности Na, K-АТФазы в составе цельных эритроцитов в условиях гипотермии.
Даларгин (Тир-D-Ала-Гли-Фен-Лей-Арг) является синтетическим аналогом нейропептида лей-энкефалинас широким спектром биологической активности. Важным эффектом пептида является антистрессорный, проявляющийся и при гипотермии [7]. Даларгин препятствует развитию стрессорной вазоконстрикции [1]. Нами показано, что предварительное введение даларгина предотвращает активацию процессов перекисного окисления липидов и снижение антиоксидантной защиты эритроцитов [10], а также стабилизирует структурно-функциональное состояние их мембран при умеренной гипотермии у крыс [9]. Учитывая тот факт, что Na, K-АТФаза весьма чувствительна как к составу липидов в микроокружении фермента, так и к их физико-химическому состоянию [13], представляет интерес изучение влияния даларгина на активность Na, K-АТФазы эритроцитов в условиях гипотермии.
Цель работы. Исследовать зависимость активности Na,К-АТФазы эритроцитов и содержания ионов Na+ и K+ в плазме крови и эритроцитах крыс от глубины гипотермии, а также возможности коррекции обнаруженных изменений путем введения даларгина.
Материалы и методы исследования. Эксперименты проведены на самцах беспородных белых крыс массой 180–200 г. Гипотермию вызывали охлаждением животных в плексигласовых камерах с рубашкой, через которую циркулировала холодная (5 °С) вода. Температуру тела крыс равномерно (0,28 °С/мин) снижали до 30 °С (кратковременная умеренная гипотермия) и 20 °С (глубокая гипотермия). За 30 мин до декапитации (контроль) или за 30 мин до начала снижения температуры тела животным внутрибрюшинно вводили 0,5 мл фармакопейного препарата даларгина в дозе 100 мкг/кг массы. После декапитации животного собирали кровь в пробирку с гепарином (50 ед/мл). Эритроциты осаждали центрифугированием при 2000 об/мин в течение 10 мин, а затем трижды промывали 0,145 М раствором NаСl, приготовленном на 0,02 М трис-НСl буфере (рН 7,6 при 20 °С).
Активность Na, К-АТФазы в цельных эритроцитах определяли по методу Казёнова А.М. и сотр. [4]. Для образования пор, проницаемых для АТФ, эритроциты обрабатывали твином-20. Преинкубацию цельных эритроцитов с твином-20 проводили в следующих условиях: отмытые эритроциты смешивали с равным объёмом 1 % твина-20, приготовленного на 0,145 М NаСl в 0,02 М трис-НСl буфере, и смесь инкубировали при комнатной температуре в течение 60 мин. Далее взвесь эритроцитов разводили в 4 раза 0,145 М NаСl в 0,02 М трис-НСl буфере без детергента. Для определения активности Na,К-АТФазы 0,2 мл конечной суспензии эритроцитов, обработанных твином-20, вводили в 1 мл инкубационной среды, содержащей (в мМ концентрациях) Na+– 100, К+– 10, трис-НСl – 50, АТФ – 3, МgCl2 – 3, ЭДТА – 1. Инкубацию проводили при 37 °С в течение 30 мин. Реакцию останавливали добавлением 0,5 мл 20 % ТХУ. Активность Na, К-АТФазы рассчитывали по разнице между АТФ-азной активностью в отсутствии и присутствии 0,2м Муабаина. Об активности фермента судили по накоплению в среде инкубации неорганического фосфора и выражали в мкмоль Рн/мл/ч.
Содержание ионов натрия и калия в плазме крови и эритроцитах определяли на атомно-абсорбционном спектрофотометре AAS-1N (CarlZeiss, Германия).
Статистическую обработку результатов осуществляли с помощью пакетов прикладных программ Statistica 6.0 и Excel общепринятыми методами вариационной статистики. В таблице результаты представлены в виде средних значений (М) ± стандартная ошибка (m). Для определения статистической значимости полученных результатов были использованы: параметрический t-критерий Стьюдента (t-тест), непараметрический U-критерий Манна – Уитни (U-тест). Достоверно различающимися признавали значения при р<0,05.
Результаты исследования и их обсуждение. Острая кратковременная гипотермия 30 °С приводит к повышению активности Na, K-АТФазы на 38 % относительно контроля (баблунок). При глубокой гипотермии активность фермента остается повышенной, хотя несколько снижается относительно умеренной гипотермии. Таким образом, независимо от длительности гипотермии активность Na, K-АТФазы эритроцитов повышается.
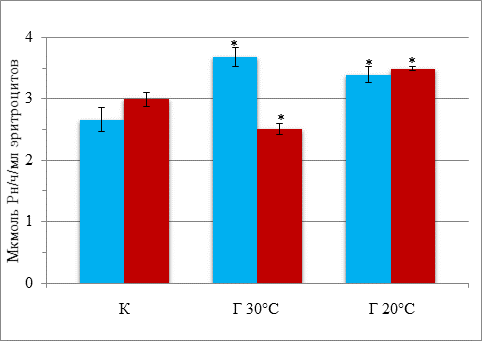
Активность Na, K-АТФазы эритроцитов при гипотермии и введении даларгина
![]() – без введения даларгина;
– без введения даларгина; ![]() – на фоне введения даларгина;
– на фоне введения даларгина;
К – контроль; Г30 °С и Г20 °С – гипотермия 30 °С и 20 °С соответственно.
* – достоверность различий относительно контроля
Ранее на отмытых от гемоглобина мембранах эритроцитов крыс нами было обнаружено снижение активности этого фермента при гипотермии 20 °С [6]. Чем же объяснить такие различия в изменении активности фермента в цельных эритроцитах и в белых тенях эритроцитов при гипотермии? Было показано, что последовательное удаление внутриклеточного содержимого и периферических белков мембран эритроцитов крыс – спектрина и актина, сопровождается прогрессивным падением активности Na, K-АТФазы [5]. Авторы показали, что как в гемолизате эритроцитов, так и в растворе смеси солюбилизированных белков мембран цитоскелета присутствуют факторы, активирующие Na, K-АТФазуэритроцитарной мембраны и головного мозга крыс, причем степень эффекта зависела от вида мембран и проявляется более выражено в случае мембранных препаратов, лишенных периферических белков. Недавно было показано, что в зависимости от способа получения теней эритроцитов и солюбилизированных белков мембранного скелета наблюдается как активирование, так и ингибирование Na, K-АТФазы удаленными с мембраны белками скелета [11]. Авторы пришли к выводу, что в смеси солюбилизированных белков мембранного скелета эритроцитов содержится фактор (факторы) Mg2+, Ca2+-зависимо модулирующие активность Na, K-АТФазы. Эти данные позволяют предположить, что внесение ЭДТА на стадии гемолиза эритроцитов и отмывания мембранных препаратов от внутриклеточного содержимого способствует удалению из препаратов мембран веществ, оказывающих модулирующее влияние на активность Na, K-АТФазы при гипотермии.
Активность и свойства транспортных АТФаз плазматических мембран в значительной степени определяются структурными особенностями липидного матрикса, в который погружены молекулы фермента [13]. Исследование структурных свойств мембран эритроцитов с помощью флуоресцентного зонда пиренане выявило изменение вязкости липидного бислоя после снижения температуры тела крыс до 30 и 20 °С [9]. В то же время выявлено зависимое от температуры тела снижение микровязкости зон белок-липидных контактов, которые представлены аннулярными липидами, составляющими микроокружение мембранных белков. Введение даларгина предотвращало изменение текучести аннулярных липидов.
В работе Хашимото и коллег [14] было показано, что уменьшение вязкости аннулярных липидов коррелирует с увеличением активности некоторых ферментов, в том числе Mg2+-АТФазы. Кроме того, в той же работе было обнаружено, что добавление докозагексаеновой жирной кислоты уменьшает вязкость аннулярных липидов сильнее, чем вязкость липидов всей мембраны в целом. Эти результаты позволяют предположить, что при гипотермии происходит избирательное накопление фосфолипидов с ненасыщенными жирнокислотными остатками в микроокружении мембранных белков. Это возможно за счет латеральной сегрегации липидов в мембране, а также за счет обмена с липидами и липопротеинами плазмы. При развитии гипотермии ускоряется гормонзависимый липолиз депонированных липидов и существенно возрастает уровень свободных жирных кислот в плазме крови [12]. В ходе снижения температуры тела в спектре жирных кислот плазмы крови крыс возрастает содержание ненасыщенных жирных кислот, что отражает участие последних в мембранных процессах, препятствующих понижению «текучести» липидов мембран клеток. Следует отметить, что введение даларгина предотвращает обусловленную холодовым стрессом секрецию гормонов и, соответственно, мобилизацию депонированных липидов [12].
Причиной активации Na, K-АТФазы при гипотермии может быть также увеличение неспецифической проницаемости мембраны эритроцитов для Na+. В этих условиях активация АТФазы является необходимым условием для восстановления ионного баланса внутри клетки. Для проверки этого предположения было исследовано содержание ионов натрия и калия в плазме крови и в эритроцитах. Изменение активности Na, K-АТФазы после гипотермии 30 °С не оказало значимого влияния на баланс одновалентных катионов в эритроцитах (таблица). Концентрация ионов Na+ иK+ во вне (ex)- и внутриклеточной (in) среде оставалось без изменений. Соответственно, отношение Naex+/Nain+ и Kex+/Kin+ совпадает у контрольных и подвергнутых гипотермии 30 °С крыс. После гипотермии 20 °С отношение Naex+/Nain+значительно снижается, а отношениеKex+/ Kin+ – существенно возрастает. Такое соотношение процессов указывает на повышение проницаемости эритроцитарных мембран для этих ионов.
Баланс одновалентных катионов в плазме крови и эритроцитах крыс в условиях гипотермии и введении даларгина (M±m; n = 6-8)
|
Состояние животного |
Na+, моль/л |
К+, моль/л |
||||
|
плазма (ex) |
эритроцит (in) |
ex/in |
плазма (ex) |
эритроцит (in) |
ex/in |
|
|
Контроль |
133,9±1,21 |
18,6±0,21 |
7,20 |
4,76±0,41 |
91,4±0,63 |
0,051 |
|
Контроль+ даларгин |
137,4±1,16 |
19,2±0,34 |
7,16 |
5,10±0,23 |
92,6±0,83 |
0,057 |
|
Гипотермия 30°С |
138,8±1,79 |
19,2±0,42 |
7,23 |
5,53±0,39 |
90,2±0,91 |
0,061 |
|
Гипотермия 30°С + даларгин |
135,1±1,65 |
18,6±0,64 |
7,26 |
4,87±0,29 |
92,6±0,92 |
0,052 |
|
Гипотермия 20°С |
117,9±2,07* |
23,7±0,49* |
4,97 |
6,46±0,26* |
76,3±1,67* |
0,084 |
|
Гипотермия 20°С + даларгин |
117,7±1,16* |
21,6±0,38* |
5,45 |
6,29±0,21* |
77,6±0,42* |
0,080 |
* – достоверные (р<0,05) относительно контроля различия.
Возникновение спазма периферических сосудов, в совокупности со сдвигом кривой диссоциации оксигемоглобина влево, приводит к тканевой гипоксии, нестабильности клеточных мембран, повышению травматизации форменных элементов крови [2]. Установлено, что даларгин способен нормализовать периферический сосудистый тонус в условиях стрессорной вазоконстрикции [1]. При этом ведущее значение в механизмах протекторного эффекта даларгина имеет увеличение уровня продукции NO, обусловленное изменением активности NO-синтаз. Можно предположить, что протекторный эффект даларгина на стрессобусловленные изменения гемодинамики имеет место при умеренной гипотермии, но не при глубокой. В определенной мере об этом свидетельствуют данные о влиянии даларгина на содержание одновалентных катионов в плазме крови и эритроцитах при гипотермии. Введение даларгина не оказало влияния на концентрацию одновалентных катионов в плазме и эритроцитах у контрольных животных и у животных с температурой тела 30 °С (таблица). При гипотермии 20 °С даларгин не предотвращает изменение соотношения Naex+/ Nain+ и Kex+/ Kin+. В крови даларгин быстро подвергается протеолизу [7], с чем, по-видимому, и связано отсутствие защитного эффекта пептида на распределение исследованных катионов вне и внутри эритроцитов после глубокой гипотермии.
Заключение
Острое кратковременное снижение температуры тела приводит к повышению активности Na,К-АТФазы эритроцитов, что является компенсаторной реакцией, направленной на поддержание ионного гомеостаза клетки, нарушенного под действием низкой температуры. После снижения температуры тела до 20 °С активность Na,К-АТФазы остается повышенной, но это не предотвращает нарушение ионного гомеостаза эритроцитов. Эти данные свидетельствуют о нарушении барьерных свойств мембран эритроцитов при глубокой гипотермии. В условиях гипотермии 30 °С введение даларгина полностью предотвращает повышение активности Na,К-АТФазы, вызываемое низкой температурой. Однако при глубокой гипотермии даларгин не предотвращает активацию Na, К-АТФазы, а также изменение ионного состава эритроцитов.
Библиографическая ссылка
Саидов М.Б., Газимагомедова М.М., Кличханов Н.К. ВЛИЯНИЕ ГИПОТЕРМИИ НА АКТИВНОСТЬ NA, K-АТФАЗЫ ЭРИТРОЦИТОВ И БАЛАНС ИОНОВ НАТРИЯ И КАЛИЯ В ПЛАЗМЕ И ЭРИТРОЦИТАХ КРЫС НА ФОНЕ ВВЕДЕНИЯ ДАЛАРГИНА // Современные проблемы науки и образования. 2016. № 3. ;URL: https://science-education.ru/en/article/view?id=24497 (дата обращения: 02.07.2026).