


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
ADDITIONAL CRITERION FOR ASSESSING THE DEGREE OF TISSUE HYPOXIA IN PATIENTS WITH PNEUMONIA

Кислородный гомеостаз организма обеспечивается, как известно, согласованным взаимодействием внешнего дыхания, аппарата кровообращения и кислородтранспортной системы (КТС) крови. Нарушение газового баланса может возникнуть при расстройстве или неполноценном функционировании любого из звеньев этой системы, в силу чего кислородная недостаточность, как типовой патологический процесс, сопровождает многие заболевания.
К числу заболеваний, приводящих к развитию гипоксии, относится пневмония – группа различных по происхождению, патогенезу и морфологическим характеристикам острое локальное инфекционно-воспалительное заболевание легких с преимущественным поражением респираторных отделов легких (альвеол, бронхиол) и внутриальвеолярной экссудацией [4]. При этом воспалительный процесс в бронхолегочной ткани вызывает значительные нарушения вентиляционной способности легких (чаще рестриктивные), что подтверждается данными спирографии.
Гипоксическое состояние, обусловленное неполноценным функционированием системы внешнего дыхания, предопределяет необходимость формирования компенсаторных перестроек, в реализации которых участвуют различные компоненты КТС организма. Важную роль в адаптации организма к дефициту кислорода играют молекулярно-генетические механизмы. В последние десятилетия было показано, что регуляция многих процессов, ответственных за транспорт кислорода осуществляется на транскрипционном уровне и модулируется гипоксия - индуцируемым фактором (hypoxia - inducible factor – HIF-1), который действует как прямой кислородный сенсор [7,8]. Он представляет собой гетеродимерный белок, кислородчувствительная субъединица которого HIF-1α, содержит пролил-гидроксилазу (ПГ), «определяющую» внутриклеточное содержание кислорода. Эта оксигеназа использует молекулярный кислород в реакциях гидроксилирования и поэтому осуществляет прямую связь между наличием молекулярного кислорода и регуляцией HIF-1. При снижении уровня кислорода в клетке происходит инактивация ПГ, что способствует стабилизации HIF-1, его переходу в ядро с последующим образованием транскрипционно-активного комплекса [10].
Мишенями HIF-1 является около 180 генов, экспрессирующих специфические белки, необходимые в условиях сниженного снабжения клеток кислородом. К ним относится в, частности, ангиотензин- конвертирующий фермент, другие модуляторы сосудистого тонуса, эритропоэтин и другие индукторы эритропоэза. Под его влиянием также усиливается экспрессия гена, контролирующего синтез γ-цепей Hb в эритробластах и возрастание в крови доли FetHb [7]. Обладая повышенным сродством к кислороду, он вносит существенный вклад в приспособление организма к новым условиям с измененной газовой средой при патологических состояниях, протекающих с гипоксией [3,5].
В этой связи представляет интерес изучение степени участия механизмов, реализующихся через активацию транскрипционного фактора HIF-1 и синтез специфических белков, в адаптации организма к недостатку кислорода при пневмонии. При этом об активизации экспрессии гена фетального гемоглобина можно судить по содержанию FetHb в периферической крови, а о состоянии эритропоэза – посредством изучения возрастного состава популяции циркулирующих эритроцитов.
Цель исследования: Оценка диагностической значимости анализа популяционного состава эритроцитов периферической крови и определения количества FetHb при пневмонии и выявление вклада этих механизмов в обеспечение адаптации КТС организма к кислородному стрессу, обусловленному патогенезом основного заболевания.
Материалы и методы исследования. Обследована группа военнослужащих (ВС) по призыву (n=30) в возрасте от 18 до 23 лет с рентгенологически установленным диагнозом "внебольничная очаговая пневмония» средней степени тяжести на 3-5 день госпитализации с клиническим и инструментальным подтверждением. Группу сравнения составили военнослужащие в возрасте 18-23 лет (n=34), находящиеся на срочной службе в рядах ВС МО РФ, признанные на момент призыва здоровыми и годными к военной службе.
Спирография проводилась на компьютерно-диагностическом комплексе "Валента" (Санкт-Петербург), определяли статические показатели: частота дыхания, форсированная жизненная емкость легких (ФЖЕЛ), динамические (объем форсированного выдоха за 1 сек.) (ОФВ1), индекс Тиффно. Гематологические исследования проводились с помощью анализатора «ADVIA 60» (Германия). В крови определялись следующие показатели: количество эритроцитов (RBC), концентрация Hb, уровень гематокрита (Нt), средний объем эритроцитов (MCV), средняя концентрация Hb в эритроците (MCHC). На аппарате «RAPIDLAB865» (Германия) измерялись рО2 и рСО2, кислородная сатурация (satO2), а также содержание оксигенированного Hb (O2Hb), FetHb, карбоксигемоглобина (COHb) и метгемоглобина (MetHb), показатели кислотно-основного состояние (КОС) крови: рН крови, содержание бикарбонатов крови (НСО3), дефицит (избыток) оснований крови (ВЕ). Рассчитывалась кислородная емкость крови (КЕК). Кинетика кислотного гемолиза изучалась по методу И.И Гительзона [1] и дополнялась построением и анализом эритрограмм. Статистическую обработку результатов проводили, используя t-критерий Стьюдента и метод факторного анализа.
Результаты исследования и их обсуждение. Исследование функции внешнего дыхания у больных с пневмонией показало, что статические и динамические показатели респираторной системы варьируют в пределах возрастной нормы, но имеют значимые различия с контролем. Так, величина ФЖЕЛ снижена у больных на 18,4±1,1%, ОФВ1 - на 15,2±1,6%, индекс Тиффно - на 14±1,0% соответственно (р≤0,05) (рис.1).
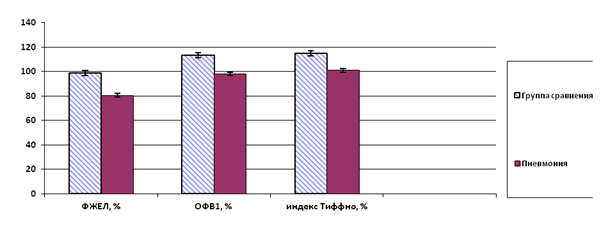
Рис.1. Показатели функции внешнего дыхания в группах обследования (достоверные отличия средних при р‹ 0,05)
Ослабление вентиляционной функции внешнего дыхания подтверждается выраженным уменьшением величины рО2 (с 82,2±2,5 мм рт ст до 65,2± 1,9 мм рт ст) и рСО2 (с 42,5± 1,8 мм рт ст до 38,1±1,5 мм рт ст) в крови обследуемых с пневмонией (р‹0,05) (рис.2). Вместе с тем, нами выявлено, что кислородная сатурация крови (satO2) достоверно не отличается в группах ВС (94,8±0,5% в группе контроля и 93,2±1,0% в группе больных). Это свидетельствует о развитии компенсации, направленной на поддержание приемлемого уровня насыщения крови O2.
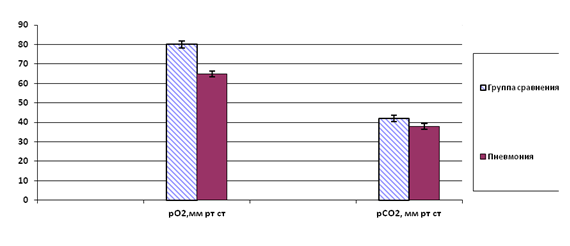
Рис.2. Показатели газового режима крови в группах обследуемых (достоверные отличия средних при р ‹ 0,05).
Возникающее вследствие нарушения газового состава крови значимое увеличение рН (с 7,40 до 7,43) (р‹0,05) и снижение стандартного бикарбоната (с 1,27 до 0,68 ммоль/л) (р‹0,05) можно расценивать как проявление частично компенсированного респираторного алкалоза [2].
Дефицит О2, обусловленный уменьшением его поглощения легкими, протекает на фоне достоверного снижения суммарных показателей красной крови (RBC, Hb, Ht) без изменения уровня индивидуальных характеристик эритроцитов (объема клеток и их насыщенность Hb) (табл.1). Это позволяет предположить, что одной из причин снижения кислородной емкости крови является уменьшение стойкости эритроцитов. Изучение кислотной резистентности эритроцитов выявило у больных пневмонией укорочение общего времени гемолиза, сдвиг максимума эритрограммы влево и увеличение самого максимума, свидетельствуя о резком падении стойкости эритроцитов.
Таблица 1
Показатели красной крови в группах обследованных
|
Показатели |
Группы обследованных военнослужащих |
|
|
Группа сравнения Группа 1 |
Пневмония Группа 2 |
|
|
Эритроциты, 1012 /л |
4,98±0,06 |
4,64 ± 0,09* |
|
Hb, г/л |
147,2±1,7 |
136,1±2,7* |
|
MCV, fl |
86,5±0,8 |
86,4±0,6 |
|
MCHC, p/dl |
34,1±0,1 |
33,9±0,2 |
|
Ht, % |
43,04±0,6 |
40,2±0,8* |
|
КЕК, мл/л |
194 ± 1,5 |
172,7 ± 1,8* |
Обозначения: * - достоверное отличие показателей в группах при р ‹ 0,05
При этом доля эритроцитов с пониженной стойкостью оказалась вдвое выше, чем аналогичных форм в группе сравнения, а количество высокостойких клеток при пневмонии, напротив, резко снижается (табл.2).
Таблица 2
Показатели кинетики кислотного гемолиза и процентное содержание эритроцитов по стойкости в группах обследованных
|
Показатели кинетики гемолиза и структуры популяции эритроцитов по стойкости |
Группа Сравнения |
Пневмония |
|
Общее время гемолиза, мин |
4,97±0,09* |
4,38±0,08* |
|
Начало гемолиза, мин |
1,97±0,06 |
1,77±0,07 |
|
Продолжительность гемолиза, мин |
2,91±0,09 |
2,51±0,08 |
|
Максимум гемолиза, мин |
3,25±0,05* |
2,70±0,07* |
|
Низкостойкие эритроциты,% |
10,0±3,5 |
7,0±3,0 |
|
Эритроциты пониженной стойкости, % |
15,5± 4,3 * |
32,0± 5,5 * |
|
Среднестойкие эритроциты, % |
62,0±6,4 |
57,0±5,8
|
|
Высокостойкие эритроциты, % |
12,5 ±3,8 * |
4,0± 2,3 * |
Обозначения: * - достоверное отличие показателей в группах при р ‹ 0,05
Очевидно, опосредованный кислородным голоданием стресс-эритропоэз сопровождается поступлением в циркуляцию функционально неполноценных эритроцитов. Они подвергаются ускоренной элиминации из сосудистого русла, что и вызывает снижение количественных показателей красной крови (содержание эритроцитов, Hb) с сохранением корпускулярных параметров (MCV, MCHC).
Одновременно при анализе отдельных фракций Hb обнаружено возрастание доли в периферической крови FetHb, (с 2,90±0,31% в контроле до 5,43±1,05% у больных) (р‹0,05), что можно расценивать как компенсацию КТС на снижение кислородной емкости крови.
Таким образом, изучение популяционного состава эритроцитов и гемоглобинового профиля позволяет допустить, что компенсаторные реакции на дефицит кислорода в клетках реализуются при пневмонии через активизацию гена эритропоэтина и фетального гемоглобина, т.е. осуществляются на транскрипционном уровне. Это заключение в целом согласуется с результатами исследования состояния крови у пациентов с серповидно-клеточной анемией (СКА) [9]. Авторами работы показано, что возрастание доли фетального гемоглобина в периферической крови больных СКА происходит на фоне сохранения среднего объема эритроцитов и ускоренного эритропоэза.
Для проверки высказанного предположения мы изучили структуру взаимосвязей учтенных в исследовании параметров КТС и КОС с помощью факторного анализа (табл.3).
Таблица 3
Факторные структуры показателей системы транспорта кислорода в группах сравнения
|
|
F1 |
|
F2 |
|
F3 |
|
|
|
контроль. |
пневмония |
контроль |
Пневмония |
контроль |
пневмония |
|
Эритроциты |
|
|
0,72 |
-0,74 |
|
|
|
Hb |
|
|
0,71 |
-0,74 |
0,61 |
|
|
Ht |
|
|
0,75 |
-0,73 |
|
|
|
MCHC |
|
|
|
|
|
-0,68 |
|
pH |
|
|
0,60 |
0,66 |
|
|
|
HCO3 |
0,80 |
|
|
|
|
-0,67 |
|
ВЕ |
0,78 |
|
|
|
|
-0,78 |
|
pCO2 |
0,62 |
|
|
-0,75 |
|
|
|
pO2 |
|
-0,78 |
|
|
|
|
|
satО2 |
|
-0,85 |
|
|
|
|
|
O2Hb |
|
-0,89 |
0,71 |
|
|
|
|
FetHb |
|
0,86 |
|
|
|
|
|
Общее время гемолиза |
|
|
|
-0,72 |
|
|
|
Дисперсия, % |
27 |
27 |
21 |
25 |
14 |
18 |
Примечание. Приведены только достоверные нагрузки переменных к фактору
В группе контроля выделено 3 фактора, два из которых являются тривиальными: F1 характеризует КОС, F2 – состояние красной крови. В структуре F2 наблюдается закономерность, описывающая эффект Бора. При нормальном функционировании системы красной крови повышение рН способствует возрастанию степени оксигенации Hb и, наоборот, при тенденции к закислению крови увеличивается отдача кислорода тканям.
В то же время в группе больных пневмонией выделено 2 равноценных по емкости фактора (F1 27% и F2 25%), участвующих в формировании адаптивного ответа на гипоксию. F1 – фактор компенсации кислородной недостаточности на молекулярно-генетическом уровне: высокие отрицательные корреляции к нему имеют рО2, satO2, O2Hb; положительную – FetHb. Снижение рО2, вызывающее нарушения гомеостаза и сопровождающееся нарушением функции внешнего дыхания, способствует экспрессии гена FetHb [6].
Другой отличительной чертой структуры взаимосвязей при пневмонии является наличие в факторе красной крови наряду с суммарными характеристиками параметра кислотного гемолиза. Одинаковые знаки этих показателей к F2 подтверждают выводы о связи снижения функциональной активности красной крови с ослаблением устойчивости эритроцитов.
Таким образом, нами продемонстрированы новые возможности традиционного и достаточно известного метода изучения возрастной популяции эритроцитов, предложенной Гительзоном в 1959 год. Параметры кислотного гемолиза имеют большой информативный потенциал для оценки степени тканевой гипоксии, что дает возможность их использования в полной мере и в наши дни наряду с современными достижениями экспериментальной науки. Можно полагать, что при рассматриваемой патологии уменьшение активности красной крови обусловлено снижением устойчивости основной популяции эритроцитов вследствие стресс-эритропоэза. Возникающий при этом кислородный дисбаланс приводит, очевидно, к усилению экспрессии гена фетального гемоглобина, которое проявляется только во время ускоренного эритропоэза [9].
Все вышесказанное позволяет заключить, что уровень FetHb в периферической крови можно использовать как индикатор гипоксического состояния, сопровождающегося снижением уровня доставки О2 тканям.
Заключение. В проведенном исследовании обнаружено, что при пневмонии функциональная несостоятельность системы кислородного обеспечения организма на уровне внешнего дыхания дополняется эритропоэтическим стрессом, снижающим резервные возможности красной крови. Нарушение кислородного баланса в тканях способствует активации экспрессии гена FetHb и возрастанию в периферической крови доли данной фракции, увеличивающей способность гемоглобина связывать кислород при относительно низком его содержании в крови. Это свидетельствует о повышении адаптационного потенциала кислородтранспортной системы крови на ранних этапах заболевания и позволяет судить о резервных возможностях, определяющих соответствие снабжения тканей кислородом их функциональной потребности. Уровень FetHb можно использовать как дополнительный критерий для установления степени тканевой гипоксии у больных пневмонией.
Рецензенты:
Ибрагимов Р.И., д.б.н., профессор, заведующий кафедрой биохимии и биотехнологии ФГБОУ ВПО «Башкирский государственный университет», г. Уфа;
Ханов А.М., д.м.н., профессор кафедры онкологии ГБОУ ВПО "Башкирский государственный медицинский университет", г. Уфа.
Библиографическая ссылка
Усманова С.Р., Шамратова В.Г., Исаева Е.Е. ДОПОЛНИТЕЛЬНЫЕ КРИТЕРИИ ОЦЕНКИ СТЕПЕНИ ТКАНЕВОЙ ГИПОКСИИ У БОЛЬНЫХ ПНЕВМОНИЕЙ // Современные проблемы науки и образования. 2014. № 6. ;URL: https://science-education.ru/en/article/view?id=16216 (дата обращения: 29.05.2026).