


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
SOME PARAMETERS DROUGHT RESISTANCE SPECIES OF GENUS SYRINGA L.GIVEN THE SHARP CONTINENTAL CLIMATE (ILLUSTRATED ORENBURG)

Введение
Одним из наиболеегубительныхабиотических факторов, действующих на растения, является недостаток влаги. Засухоустойчивость – это свойство растений переносить перегрев и обезвоживание, а также в ходе онтогенеза приспосабливаться к действию засухи;осуществлять в данных условиях нормальный рост, развитие и воспроизведение благодаря наличию ряда свойств, возникающих в процессе эволюционного развития под влиянием условий существования. Способность растений приспосабливаться к условиям окружающей среды является результатом изменчивости, наследственности или отбора. Жаростойкость, засухоустойчивость сформировались в ходе эволюции в результате действия высоких температур, засухи [3].
Характеристика климатических условий района исследования
Климат Оренбургской области резко-континентальный, т.е. для него характерна морозная зима с устойчивым снежным покровом и жаркое лето с малым количеством осадков, а также высокие годовые амплитуды температуры. Среднегодовая сумма активных температур составляет 2400–2600°С, продолжительность вегетационного периода – 150– 155 дней, абсолютный максимум положительных температур может достигать +41°С. Сумма температур выше +5°С по г. Оренбургу равна 2300–2400°С [10].
По данным о температуре за время вегетации, предоставленным Оренбургским областным центром по гидрометеорологии и мониторингу окружающей среды, нами был рассчитан гидротермический коэффициент (ГТК) Г.Т. Селянинова[8], который является показателем обеспечения определенной территории влагой.
Проанализировав данные о температурных условиях девяти вегетационных периодов (таб. 1), было установлено, что для г. Оренбурга характерен ГТКср. = 0.3, что позволяет отнести территорию города к сухой зоне (ГТК<0,4). Это связано с неустойчивостью увлажнения, а также с неравномерностью выпадения осадков.
Таблица 1
Климатические условия периодов вегетации растений за период с 2009 по 2013 г. по г. Оренбургу
|
Климатические параметры |
2009 |
2010 |
2011 |
2012 |
2013 |
|
|
Температура, °С |
средн. |
+19,1 |
+22,2 |
+19,1 |
+21,3 |
+19,4 |
|
min |
0,0 |
+3,3 |
+1,4 |
+1,5 |
+2,3 |
|
|
max |
+35,4 |
+37,8 |
+39,9 |
+38,6 |
+35,6 |
|
|
Относительная влажность воздуха, % |
средн. |
54 |
40 |
57 |
51 |
55 |
|
min |
13 |
8 |
12 |
11 |
10 |
|
|
Скорость ветра, м/сек |
средн. |
2,9 |
3,1 |
2,9 |
3,1 |
2,9 |
|
max |
11 |
10 |
11 |
12 |
11 |
|
|
Осадки |
сумма, мм |
77 |
30 |
67 |
64 |
235 |
|
мax за 12 ч, мм |
19,0 |
5,0 |
21,0 |
24,0 |
44,0 |
|
|
число дней с осадками |
33 |
24 |
43 |
29 |
68 |
|
Материал и методика исследований
В качестве объектов исследования были выбраны 7 видов сирени (таб. 2):
Таблица 2
Характеристика изучаемых видов сирени
|
Вид |
Секция |
Ареал естественного распространения |
Климатические условия естественного местообитания |
|
S. vulgarisL. |
Vulgaris C.K. Schneid. |
Югославия, Румыния, Болгария, на незначительной части Турции |
климат умеренно-континентальный. tср самого жаркого месяца 22–24 °С, а самого холодного –-2-5 °С. Осадки: 500 – 700 мм/год |
|
S. pubescensTurkz. |
СеверныйКитай |
различные климатические условия, т.к. эта область занимает огромную гористую территорию. Резко отличаются между собой климат Приамурья и Приморья Дальнего Востока, западного Китая, Корейский полуостров и Япония. |
|
|
S. persicaL. |
Карпаты |
климат – континентальный. tср января 6.5 °С, июля — 18,5 °С. Осадки: 1500–1700 мм/год |
|
|
S. х Henry C.K. Schneid. |
VillosaeC.K. Schneid.
|
Франция |
различные климатические условия, т.к. эта область занимает огромную гористую территорию. Резко различается между собой климат Приамурья и Приморья, Дальнего Востока, западного Китая, Корейский полуостров и Япония. |
|
S. KomarowiiC.K. Schneid. |
юго - западныйКитай |
||
|
S. SweginzowiiKoehne |
Китай |
||
|
S. josikaeaJacq. |
Карпаты |
климат умеренно-континентальный. tср самого жаркого месяца 22–24 °С, а самого холодного –- 2–5 °С. Осадки 500–700 мм/год |
Определение водного дефицита (ВД)производилось по «Программе и мотодике…» [6] по следующей формуле:
ВД =![]() х 100%, гдеА – масса сухой навески;Б
– масса воды перед насыщением;В – масса воды после полного насыщения;Г – масса листьев
после полного насыщения водой.
х 100%, гдеА – масса сухой навески;Б
– масса воды перед насыщением;В – масса воды после полного насыщения;Г – масса листьев
после полного насыщения водой.
Также определяли: общую оводненность листьев (W)и водоудерживающую способность (R) – по методике Н.А. Гусева [4], содержание«подвижной влаги»(L) в пробах по методике В.А. Таренкова, Л.Н. Ивановой [9] по следующим формулам:
W= ![]()
R=![]()
L= W- R, гдеМ – масса свежей пробы;М1 – масса пробы спустя сутки;М2 – масса пробы после полного высушивания.
Среднюю дифференциальную скорость водопотери определяли по методике В.И. Авдеева [1].
Результаты и обсуждение
В результате анализа литературных данных [2,5] было установлено, что засухоустойчивость сиреней определяется не только биологическими особенностями конкретного вида или сорта, но и возрастным состоянием растений. Наибольшей засухоустойчивостью обладают молодые растения сирени. С возрастом способность переносить засуху резко снижается. Стоит отметить, что избыток влаги также переносится сиренью очень плохо.Исключение в данном случае – S. josikaeaJacq., вид сирени очень требовательный к условиям высокого увлажнения. В целом все виды сирени достаточно хорошо переносят условия недостаточного увлажнения.
В результате анализа таблиц 1 и 2 становится очевидным тот факт, что климатические условия естественных местообитаний видовых сиреней заметно контрастируют с гидротермическим режимом Оренбургской области, поэтому является целесообразным проведение серии опытов для выявления физиологических особенностей засухоустойчивости вышеуказанных видов сирени в условиях нашего региона.
Изучение засухоустойчивости проводили методом искусственного завядания. Для этого отбирали пробы по 3–5 листьев в каждой в средней части куста с южной стороны растения, снабжали их этикеткой и, уложив в слегка увлажненный пакет, доставляли в лабораторию, где их погружали черешками в колбы с водой. Затем колбы помещали в кристаллизатор в целях предотвращения испарения воды на 24 часа до полного насыщения листьев водой. При этом определяли вес только что собранных листьев в полевых условиях; массу листьев после 24-часового насыщения. Далее в течение нескольких часов листья выдерживали в сушильном шкафу при температуре 105°С до абсолютно сухой массы. Все взвешивания производили на электронных лабораторных весах марки HiglandHCB 1502. Вес навесок при сборе, после полного насыщения и сухих листьев отражен в таблице 3.
Таблица 3
Характеристика массы листьев
|
Вид |
Масса сырой навески, г |
Масса листьев после полного насыщения, г |
Масса сухой навески, г |
|
S. vulgarisL. |
3,65 |
4,25 |
1,4 |
|
S. persicaL. |
1,55 |
1,7 |
0,55 |
|
S. pubescensTurkz. |
1,1 |
1,3 |
0,25 |
|
S. josikaeaJacq. |
2,9 |
3,45 |
0,95 |
|
S. sweginzowiiKoehne. |
1,9 |
2,3 |
0,65 |
|
S. komarowiiC.K. Schneid. |
1,75 |
2,1 |
0,65 |
|
S. х henry C.K. Schneid. |
1,15 |
1,3 |
0,35 |
Высокий уровень оводненности листьев в условиях водного стресса свидетельствует о повышенной способности вида растения адаптироваться к меняющимся условиям водоснабжения, о его более высокой засухоустойчивости. В данном случае наибольшим процентом оводненности обладает S. persicaL. и S. х henryC.K. Schneid.(64,5% и 61,1% соответственно), наименьшим – S. vulgarisL. – 39,7%, остальные изученные виды имеют средние значения по данному показателю.S. vulgarisL. –широко распространенныйвид сирени на территории Оренбургской области, следовательно, можно сделать вывод о том, что данный вид сирени обладает высокой экологической пластичностью и хорошо адаптировался к условиям резко-континентального климата области.Поэтому именно S. vulgarisL. использовалась нами как контрольный образец по всем показателям засухоустойчивости.
По показателю общейоводненности листа невозможно полностью охарактеризовать состояние водного режима растения определенного вида, поэтому в целях комплексной оценки степени засухоустойчивости нами дополнительно определялись водоудерживающая способность и водный дефицит листьев сирени.
Наряду с низкой оводненностью клеток и тканей листьев S. vulgarisL. показатель водоудерживающей способности у данного вида – самый высокий из всех изученных видов и составляет 22%. Это свидетельствует о том, что за счет высокого процента водоудерживающей способности компенсируется низкий уровень оводненности клеток и тканей. Минимальный процент водоудерживающей способности характерен для S. persicaL. и S. х henryC.K. Schneid., хотя уровень оводненности у данных видов гораздо выше, в отличии отS. vulgarisL.Следовательно, можно сделать вывод о том, что эти два вида сирени, содержащие большое количество воды в клетках, в условиях искусственного завядания очень быстро ее расходуют, что свидетельствует о более низком уровне засухоустойчивости данных видов среди всех изученных.
Водный дефицит– недостаток насыщения водой растительных клеток, возникающий в результате интенсивной потери воды растением, не восполняемой поглощением её из почвы. Водный дефицит обычно наблюдается в наиболее жаркие часы дня. Этот показатель объединяет в себе потенциальную способность тканей к водонасыщению и реальный уровень их оводненности.
Недостаток воды из всех изученных нами видов сирени в большей степени проявляется у S. josikaeaJacq., S. sweginzowiiKoehne. и S. komarowiiC.K. Schneid., но за счет хороших показателей, проявляемых данными видами по общей оводненности и водоудерживающей способности листьев, можно сделать вывод о том, что эти виды сиреней переносят условия недостаточного увлажнения или даже засухи без существенных изменений в их жизненном состоянии (таб. 4). Кроме этого, водный дефицит в природной среде – величина достаточно изменчивая и зависит от конкретных условий водоснабжения или погоды в течение суток [3].
Таблица 4
Характеристика показателей засухоустойчивости видов сирени
|
Вид |
Оводненность тканей (W,%) |
Водоудерживающая способность (R,%) |
ВД, % |
|
39,7 |
22 |
21 |
|
|
S. persicaL. |
64,5 |
3,2 |
12,5 |
|
S. pubescensTurkz. |
59,1 |
18,2 |
19 |
|
S. josikaeaJacq. |
48,9 |
17,8 |
25 |
|
S. sweginzowiiKoehne. |
55,3 |
10,5 |
24,2 |
|
S. komarowiiC.K. Schneid. |
54,3 |
8,6 |
24,1 |
|
S. х henry C.K. Schneid. |
61,1 |
5,56 |
20 |
Изучая содержание подвижной влаги и уровень водопотери как показателей засухоустойчивости, было установлено, что показатель водоудерживающей способности клетки обратно пропорционален показателю потери воды. Таким образом, наибольшей степенью засухоустойчивости по параметру потери воды обладает S. vulgarisL., наименьшей – S. persicaL. и S. х henryC.K. Schneid. Остальные изученные виды сирени имеют более средние значения. Кроме этого, для S. persicaL. и S. х henryC.K. Schneid. характерен высокий процент содержания подвижной влаги в клетках, в отличие от S. vulgarisL., что свидетельствует о проявлении низкого уровня засухоустойчивости этих двух видов по сравнению со всеми изученными.
Рис. 1
Оценка засухоустойчивости видов сирени по содержанию подвижной влаги и количеству потерянной воды (за сутки)
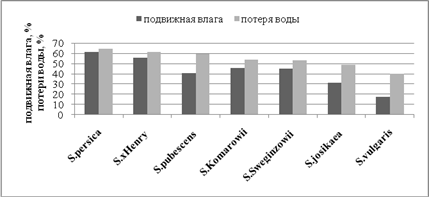
Среднюю дифференциальную скорость водопотери рассчитывали следующим образом. На каждом этапе последовательных взвешиваний (n) через интервалы времени t = 1, 2, 4, 6, 8, 12, 16, 20, 22, 24 ч после начала опыта (t = 0) определяли массу потерянной воды (мг за 1 ч), для чего из значения сырой массы листа каждого этапа (Р0, Р1, Р2, Р4 и т.д.) вычитали Р последующего этапа [например, (Р0 – Р1) /t, (Р1 – Р2) /t, (Р2 – Р4) /t и т.д.]. В итоге получали дифференциальную скорость водопотери (ДСВ) на каждом этапе завядания. После этого вычисляли среднее значениеСДСВср. = ∑(ДСВ)/n (мг за 1 ч), затем, какобычно, относили СДСВср. к исходной сырой массе листьев при полном насыщении (СМПН, г), т.е. СДСВср./СМПН (мг/г за 1 ч). Предложенный показатель СДСВ учитывает не только относительную массу испарившейся воды в процессе завядания, но и длительность завядания, поскольку одна и та же величина водопотери может быть достигнута, например, за 4 ч и за 12 ч. В последнем случае расчётная величина СДСВ минимальная.
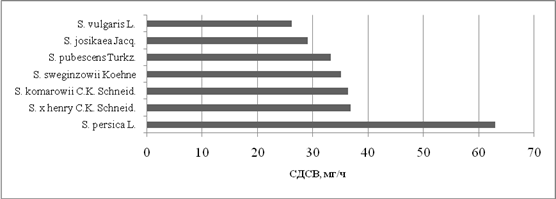
Среди всех изученных видов сирени наибольшей скоростьюводопотери обладает S. persicaL. (62,94 мг/ч). Это объясняется тем, что у этого вида самый высокий уровень оводненности и содержания подвижной влаги в клетках, поэтому за 24 часа листья сирени персидской потеряли достаточно большое количество воды. Что же касается S. vulgarisL., то у данного вида скорость водопотериминимальная и равна 26,2 мг/ч. В целом, рассматривая эти два вида в контексте показателя СДСВ, можно сделать вывод о том, что уровень засухоустойчивости S. vulgarisL. и S. persicaL. примерно одинаков. Низкую скорость водопотери S.vulgarisL. можно объяснить низким уровнем оводненности и содержания подвижной влаги в клетках данного вида сирени. У S. persicaL. наблюдается обратный эффект – в клетках данного растения содержится очень много свободной воды, потеря которой не наносит никакого вреда растению. Все остальные виды сирени имеют средний показатель СДСВ, варьирующий в пределах от 29 до 36,8 мг/ч. Кроме этого, целесообразно отметить, что в течение 24 часов ни один из изученных нами видов сирени не потерял весь запас воды.
Рис. 2
СДСВ листьев видовой сирени (за 24 ч)
Выводы
S. persicaL. и S. х henryC.K. Schneid. обладают довольно высокойстепенью засухоустойчивости за счет хорошей оводненностиклеток и тканей, в отличие отS. vulgarisL. и S. josikaeaJacq., имеющих хорошуюводоудерживающую способностьи соответственно низкую СДСВ.Кроме этого, из таблицы 1 видно, что S. persicaL. и S. vulgarisL. относятся к секции Vulgaris, а S. х henryC.K. Schneid. и S. josikaeaJacq. к секции Villosae. Из всего вышеперечисленного можно сделать вывод о том, что виды сирени, принадлежащие к разным таксономическим единицам, проявляют сходные физиологические показатели по параметру засухоустойчивости.Соответственно, это является одним из доказательств родства данных видов, что позволяет отнести их к одному родуSyringaL. Что же касается систематического деления видов внутри одного рода, то можно предположить, что данное деление является относительным и производится по комплексу отдельно взятых признаков, характерных для определенного вида.
В целом изучение особенностей водного режима видов родаSyringaL.позволяет сделать вывод о том, что все они проявляют достаточно высокий уровень засухоустойчивости.
Рецензенты:Сафонов М.А., д.б.н., доцент, заведующий кафедрой общей биологии, экологии и методики обучения биологии ФГБОУ ВПО «Оренбургский государственный педагогический университет», г.Оренбург.
Паршина Т.Ю.,д.б.н., доцент, профессор кафедры зоологии и физиологии человека и животных ФГБОУ ВПО «Оренбургский государственный педагогический университет», г.Оренбург.
Библиографическая ссылка
Назарова Н.М. НЕКОТОРЫЕ ПОКАЗАТЕЛИ ЗАСУХОУСТОЙЧИВОСТИ ВИДОВ РОДА SYRINGAL. В УСЛОВИЯХ РЕЗКО-КОНТИНЕНТАЛЬНОГО КЛИМАТА (НА ПРИМЕРЕ Г. ОРЕНБУРГА) // Современные проблемы науки и образования. 2014. № 2. ;URL: https://science-education.ru/en/article/view?id=12491 (дата обращения: 29.06.2026).