


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
PECULIARITIES OF GROWTH AND PRODUCTION PROCESSES OF OATS AT THE INOCULATION OF TECHNOGENIC ELUVIALS BY SOIL MICROORGANISMS

По оценкам специалистов площадь нарушенных земель в Кузбассе составляет не менее 91,7 тыс. га, включая и зоны подтопления в районах закрытых угольных шахт [10]. Добыча угля сопровождается огромным экологическим ущербом природным экосистемам. Неизбежным следствием горно-промышленного производства являются карьерные выемки, отходы в виде отвалов вскрышных горных пород, подработанные шахтовые земли. В результате нарушения почвенного и растительного покровов происходит замена природных ландшафтов техногенными, восстановление которых естественным путем идет очень медленно.
Известно, что основным источником поступления элементов зольной пищи в растения и почвенные микроорганизмы являются минералы почвообразующих пород. Наиболее четко такая зависимость проявляется в регенерационных биогеоценозах техногенных ландшафтов, характеризующиеся низким содержанием доступных растениям элементов питания [1]. Внесение инокулята почвенных микроорганизмов повышает биологическую активность техногенных элювиев и позволяет ускорить процессы первичного почвообразования на отвалах Кедровского угольного разреза, что, в свою очередь, способствует созданию благоприятных условий для роста и развития растений.
В связи с вышесказанным, целью наших исследований являлось изучение особенностей ростовых и продукционных процессов овса при инокуляции почвенными микроорганизмами в условиях породных отвалов. Исходя из поставленной цели, решались следующие задачи:
- изучить интенсивность фотосинтеза на уровне листового аппарата;
- провести анализ основных показателей роста и развития – сухой массы растений, величины площади листьев и динамики ее формирования;
- изучить структуру урожая овса.
Объекты и методы
Модельный эксперимент заложен в 2008 г. на породном отвале «Южный», на техногенных элювиях, лишенных растительности. Возраст отвала 25–30 лет, в 2004 г. проведен комплекс работ по его планировке. Породы отвала представлены песчаником (60 %), алевролитами (20 %), аргиллитами (15 %), суглинками и глинами (5 %). Преобладающей фракцией являются крупные агрегаты (от 3 до 10 и более мм), содержание мелких частиц снижено. Исследуемые элювии характеризуются щелочной реакцией (рН 7,8), низкой обеспеченностью подвижными формами фосфора и азота (7 и 1,7 мг/кг соответственно), содержание обменного калия чуть ниже нормы (125 мг/кг). Анализ содержания тяжелых металлов не показал превышение ПДК.
На основе литературных данных [4, 6, 7] для инокуляции породных отвалов были выбраны следующие эколого-трофические группы микроорганизмов – микроорганизмы, использующие минеральные формы азота, микроорганизмы, разлагающие силикаты и микроскопические грибы.
Микроорганизмы выделяли из зональных почв на специальных агаризованных средах: на среде Сабуро – микроскопические грибы, на крахмало-аммиачном агаре (КАА) – микроорганизмы, использующие минеральный азот, на среде Александрова-Зака – микроорганизмы, разрушающие силикаты. Микроскопирование полученных инокулятов показало, что микроорганизмы, разрушающие силикаты, представлены в основном бактериями родов Bacillus, Mycobacterium, Pseudomonas. На среде Сабуро преимущественно развивались микроскопические грибы родов Mucor, Penicillium, Aspergillus, Cladosporium, Trichoderma, Dematium. На крахмало-аммиачном агаре выявлено преобладание бактерий родов Actinomyces, Mycobacterium, Streptomices, Streptoverticillium, Nocardia, Micromonospora, Streptosporangium.
Инокулят получали путем наращивания микробной массы в соответствующих жидких питательных средах последовательным пересевом в возрастающие объемы среды. Испытуемые смеси микроорганизмов для инокуляции составляли в равных объемах. Внесение инокуллята микроорганизмов проводили дважды за вегетацию – 20 июня и 20 июля 2008–2010 гг.
Схема закладки пробных площадок представлена в таблице 1. Повторность опыта каждой пробной площадки 3-х кратная. Делянки по повторностям пространственно отдалены друг от друга для исключения влияния неоднородности элювиального субстрата и рельефа местности.
Таблица 1. Схема закладки опыта
|
№ ПП |
Вносимые микроорганизмы |
|
ПП 1 |
Контроль (полив водой) |
|
ПП 2 |
Микроскопические грибы |
|
ПП 3 |
Микроорганизмы, разлагающие силикаты |
|
ПП 4 |
Микроорганизмы, использующие минеральные формы азота |
|
ПП 5 |
Микроскопические грибы + микроорганизмы, разлагающие силикаты |
|
ПП 6 |
Микроскопические грибы + микроорганизмы, использующие минеральные формы азота |
|
ПП 7 |
Микроорганизмы, разлагающие силикаты + микроорганизмы, использующие минеральные формы азота |
|
ПП 8 |
Микроорганизмы, разлагающие силикаты + микроскопические грибы + микроорганизмы, использующие минеральные формы азота. |
Оценку фотосинтетической способности листьев проводили по уровню восстановленных ассимилятов бескамерным методом, разработанным О. Д. Быковым [2]. Метод основан на определении относительного изменения в удельном содержании восстановленных веществ за единицу времени. Изучение ростовых и продукционных процессов на уровне целого растения проводили по методике И. В. Кармановой [3] на основе определения массы растений и площади листьев. Анализ структуры урожая проведен по следующим показателям: количество стеблей на 1 растении, количество зерен с 1 растения, масса 1000 зерен, масса зерен с 1 растения [8].
Результаты и их обсуждение
Фотосинтетическая способность может служить диагностическим признаком состояния растений, обусловливает их рост и развитие. Метод оценки изменения восстановленных веществ в листьях овса за единицу времени позволяет судить о потенциальной способности растений к фотосинтезу.
В ходе анализа результатов за два года было установлено, что интенсивность синтеза ассимилятов при внесении микроорганизмов выше контроля (рис. 1). Оценка динамики данного процесса за два года показала, что максимальная интенсивность фотосинтеза на всех ПП наблюдалась в период с 5 по 25 июля и резко снижалась к 5 августа. В начале периода исследований (15 июня) значения уровня синтеза ассимилятов изменяются в пределах от 19,19 до 22,44 мг/г·ч; максимальные достоверные отличия отмечены на ПП 3, куда вносили микроорганизмы, разлагающие силикаты (37 %). К 25 июня максимальная интенсивность фотосинтеза наблюдалась на той же ПП и превышает контроль на 21 %. Высокие значения данного показателя также отмечены на ПП 6–8.
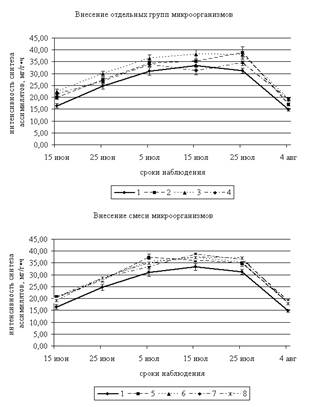
Примечание: 1 – Контроль; 2 – Грибы; 3 – Микроорганизмы, разлагающие силикаты; 4 – Микроорганизмы, использующие минеральные формы азота; 5 – Грибы + микроорганизмы, разлагающие силикаты; 6 – Грибы + микроорганизмы, использующие минеральные формы азота; 7 – Микроорганизмы, разлагающие силикаты + Микроорганизмы, использующие минеральные формы азота; 8 – Микроорганизмы, разлагающие силикаты + грибы + микроорганизмы, использующие минеральные формы азот.
Рисунок 1. Фотосинтетическая способность листьев овса, мг/г·ч (2009–2010 гг.)Максимальная фотосинтетическая активность отмечена 5 июля. Наибольшие отличия от контроля наблюдаются на ПП 5 (смесь микроорганизмов, разлагающих силикаты и микроскопических грибов) и составляют 21 %. К 15 июля интенсивность фотосинтеза остается примерно на том же уровне, однако, максимальные значения наблюдаются на ПП 3 и 7 (внесение микроорганизмов, разлагающих силикаты отдельно и в комплексе с микроорганизмами, использующими минеральный азот, соответственно), а отличие от контроля составляет 15 %. При этом на ПП 4 данный показатель ниже контроля и составляет 31,24 мг/г·ч.
Наиболее интенсивный синтез ассимиллятов в листьях овса 25 июля отмечен на ПП 2, 3 и 8 (отличие от контроля 21–24 %). К концу периода исследования (5 августа) фотосинтетическая активность на всех ПП существенно снижается и изменяется в пределах 14,8 – 19,33 мг/г·ч (минимальные значения отмечены в контроле). Однако при внесении микроорганизмов, разлагающих силикаты (ПП 3), значения данного показателя превышают контроль на 31 %. На ПП 4–7 также отмечены достаточно высокие значения интенсивности фотосинтеза.
Рост и развитие являются важнейшими характеристиками жизнедеятельности растений [5, 9]. В качестве основных показателей роста и развития, определяющих урожайность овса, могут быть – сухая масса растений, величина площади листьев и динамика ее формирования. Учет общей наземной массы позволяет определить особенности роста и продукционной деятельности в течение вегетационного периода у овса на уровне целого растения. Формирование наибольшей биомассы листьев является результатом высокой фотосинтетической активности. Ее определение на протяжении всей вегетации позволяет прогнозировать урожай растений.
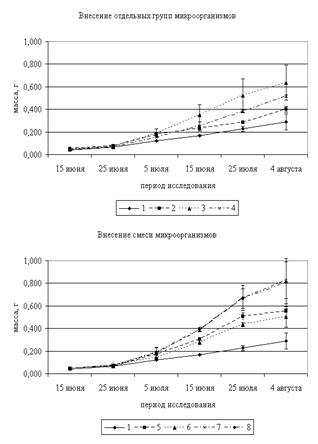
Полученные экспериментальные данные показали, что прирост сухой массы овса в вегетационный период происходит по S-образной кривой Сакса как в контрольных, так и в опытных вариантах – с минимальным приростом в июне и периодом максимального роста в конце июля – начале августа (рис. 2). Однако прирост массы растений в опытных вариантах выше во все сроки наблюдений по сравнению с контролем.
В начале периода вегетации (15–25 июня) отмечены минимальные отличия массы опытных растений от контрольных. Максимальные отличия от контроля наблюдались при внесении микроскопических грибов и составляли 19 и 9 %, 15 и 25 июня соответственно. Сухая масса опытных растений 5 июля изменяется в пределах от 0,143 до 0,189 г (в контроле 0,122 г). Максимальная масса овса наблюдаеся на ПП 3, 7 и 8 (отличия от контроля составляют 50–55 %). В последующие периоды исследования наибольшая масса овса наблюдалась на ПП 7 и 8, превышая контроль в 2,3–2,9 раз. При этом отличия от контроля для данных ПП являются достоверными.
К важным показателям, определяющим урожайность растений, относится величина площади листьев и динамика ее формирования. Формирование наибольшей биомассы листьев является результатом высокой фотосинтетической активности. Ее учет на протяжении вегетации позволяет прогнозировать урожай растений [3].
|
Рисунок 2. Динамика накопления сухой массы овса (2009–2010 гг.)
|
Рисунок 3. Динамика площади листьев овса, см2 (2009–2010 гг.) |
|
Примечание: 1 – Контроль; 2 – Грибы; 3 – Микроорганизмы, разлагающие силикаты; 4 – Микроорганизмы, использующие минеральные формы азота; 5 – Грибы + микроорганизмы, разлагающие силикаты; 6 – Грибы + микроорганизмы, использующие минеральные формы азота; 7 – Микроорганизмы, разлагающие силикаты + Микроорганизмы, использующие минеральные формы азота; 8 – Микроорганизмы, разлагающие силикаты + грибы + микроорганизмы, использующие минеральные формы азота.
|
|
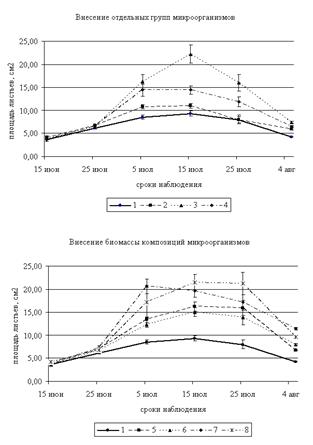
Анализ полученных результатов за 2009–2010 гг. показал, что площадь листьев овса в начале вегетации (15–25 июня) при инокуляции микроорганизмами существенно не отличается от контроля (рис. 3). Наибольшие различия отмечены в середине вегетационного периода. Максимальная площадь листьев 15 июня наблюдалась при внесении микроскопических грибов (ПП 2) и смеси всех микроорганизмов (ПП 8) и превышает контроль на 11,7 и 15,2 %, соответственно. К 25 июня площадь листьев на всех ПП увеличивается и изменяется в пределах от 6,21 (контроль) до 7,29 см2. Наибольшие значения данного показателя отмечены на ПП 5 и 7 (выше контроля на 14 и 17,4 %, соответственно).
Максимальные значения площади листьев 5 июля наблюдались на ПП 7 (внесение комплекса микроорганизмов, разлагающих силикаты и использующих минеральный азот) и составляли 20,66±1,56 см2, что выше контроля в 2,4 раза. Достаточно высокие значения отмечены также при внесении микроорганизмов, разлагающих силикаты (выше контроля 1,9 раз) и комплекса всех микроорганизмов (выше контроля в 2 раза). Максимальная площадь листьев за вегетацию наблюдалась 15 июля. В данный период исследования наибольшие показатели отмечены на тех же пробных площадках: на ПП 3 – выше контроля в 2,4 раза, на ПП 7 – в 2,1 раз, на ПП 8 – 2,3 раза.
К 25 июля площадь листьев начинает уменьшаться в результате старения и отмирания листьев. Наибольшая площадь листьев в данный период наблюдается на ПП 3, 7 и 8 (выше контроля в 2, 2,1 и 2,7 раз, соответственно). Кроме того, высокие значения отмечены при внесении смеси грибов с микроорганизмами, разлагающими силикаты (ПП 5) – выше контроля в 2 раза. В конце периода исследования максимальные значения площади листьев отмечены на ПП 7 и 8, отличия от контроля составляют 170 и 128 %, соответственно. Достоверные отличия площади листьев от контроля наблюдались 5 и 25 июля, 4 августа.
К концу роста и развития растений результатом их фотосинтетической деятельности через цепь превращения веществ и энергии является определенный потенциал продуктивности, мерой которого считается урожай – масса 1000 зерен, количество зерен в колосе, масса зерна с растения [8].
Анализ экспериментальных данных показал, что внесение микроорганизмов способствует увеличению основных показателей структуры урожая (табл. 2).
Таблица 2. Структура урожая овса (2009–2010 гг.)
|
№ ПП |
Количество зерен с одного растения |
Масса зерен с одного растения, г |
Масса 1000 зерен, г |
|
1 |
2,09±0,09 |
0,04±0,005 |
14,06±0,62 |
|
2 |
3,90±0,30* |
0,12±0,005* |
24,41±0,91* |
|
3 |
5,71±0,43* |
0,20±0,010* |
30,36±0,93* |
|
4 |
3,82±0,39* |
0,10±0,005* |
25,38±0,75* |
|
5 |
3,21±0,31* |
0,09±0,005* |
26,63±0,13* |
|
6 |
3,71±0,08* |
0,09±0,005* |
25,41±0,03* |
|
7 |
4,47±0,04* |
0,12±0,010* |
24,53±0,65* |
|
8 |
6,03±0,08* |
0,18±0,005* |
27,72±1,60* |
Примечание: * отмечены достоверные отличия от контроля при р<0,05.
1 – Контроль (полив водой); 2 – Грибы; 3 – Микроорганизмы, разлагающие силикаты; 4 – Микроорганизмы, использующие минеральные формы азота; 5 – Грибы + микроорганизмы, разлагающие силикаты; 6 – Грибы + микроорганизмы, использующие минеральные формы азота; 7 – Микроорганизмы, разлагающие силикаты + Микроорганизмы, использующие минеральные формы азота; 8 – Микроорганизмы, разлагающие силикаты + грибы + микроорганизмы, использующие минеральные формы азота.
Количество зерен с одного растения превышает контроль в 1,5–2,9 раз, масса зерен с одного растения – в 2,3–5 раз, масса 1000 зерен – в 1,7–2,2 раза. При этом все отличия достоверны. Максимальные показатели структуры урожая отмечены на ПП 3 и 8. Таким образом, внесение микроорганизмов, разлагающих силикаты отдельно и в смеси с другими группами микроорганизмов, способствует увеличению урожая овса.
Заключение
Анализ полученных данных показал, что наибольшему увеличению сухой массы, площади листьев и структуры урожая овса способствует внесение микроорганизмов, разлагающих силикаты как отдельно (ПП 3), так и в смеси с микроорганизмами, использующими минеральный азот (ПП 7) и композиции из всех микроорганизмов (ПП 8). Наибольшие значения интенсивности фотосинтеза в течение вегетации наблюдаются при инокуляции микроорганизмами, разлагающими силикаты (ПП 3).
Рецензенты:
Еремеева Н.И., д.б.н., профессор, профессор кафедры зоологии и экологии ГОУ ВПО Кемеровский государственный университет, г. Кемерово.
Куприянов А.Н., д.б.н., профессор, заведующий отделом «Кузбасский ботанический сад» ГБУН Института экологии человека, г. Кемерово.
Библиографическая ссылка
Корниясова Н.А., Неверова О.А. ОСОБЕННОСТИ РОСТОВЫХ И ПРОДУКЦИОННЫХ ПРОЦЕССОВ ОВСА ПРИ ИНОКУЛЯЦИИ ТЕХНОГЕННЫХ ЭЛЮВИЕВ ПОЧВЕННЫМИ МИКРООРГАНИЗМАМИ // Современные проблемы науки и образования. 2013. № 6. ;URL: https://science-education.ru/en/article/view?id=11607 (дата обращения: 17.07.2026).