


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
EPIGENETIC VARIABILITY OF NON-METRIC SIGNS OF ANIMALS FROM POPULATIONS WITH DIFFERENT DEGREES OF ANTHROPOLOGICAL LOADING

Введение
Возрастание значимости и развитие методологии комплексного экологического мониторинга в последние три десятилетия способствовало повышению интереса к изучению эпигенетической изменчивости в популяциях разных видов животных и растений [2; 5-9]. В ряде независимых исследований установлена взаимосвязь между «благополучием» условий среды и стабильностью популяционного гомеореза, наилучшим индикатором которой служит уровень флуктуирующей асимметрии [1; 6]. Высокая стабильность развития поддерживается на базе генетической коадаптации при оптимальных условиях развития [10]. В связи с этим стабильность эпигенеза может использоваться как мера средового стресса в мониторинге здоровья среды [6].
Цель данной работы заключалась в изучении реализованной эпигенетической изменчивости и стабильности гомеореза в популяциях животных в градиенте действия антропогенных факторов. Объектами исследования явились клоп-солдатик (Insecta, Hemiptera, PyrrhocorisapterusL., 1758) и узкочерепная полёвка (Mammalia, Rodentia, MicrotusgregalisPall., 1779), распространённые и многочисленные в районе исследования, что позволило собрать репрезентативный материал.
Материалы и методы исследования
У клопа-солдатика анализировали четыре элемента рисунка надкрылий: чёрное верхнее пятно (А) и нижнее (Д), чёрное верхнее окаймление на красном фоне надкрылий (В) и нижнее (Т) (рис.1). Проанализировали 100 экземпляров клопов из двух локальных популяций (лесопарк на окраине города и район города с высокой антропогенной нагрузкой) по 50 единиц в каждой. Городская популяция расположена в районе мясокомбината города Ишима. Рядом находятся три автозаправочные станции, железнодорожное полотно транссибирской магистрали и пересечение автомагистралей. Сбор материала проводили одновременно в разных местообитаниях в конце мая 2013 года. Достоверность различий между фенотипическими классами внутри каждой популяции и между популяциями определяли с помощью критерия χ2 К. Пирсона.

Рис.1. Структура рисунка надкрылий клопа-солдатика
Узкочерепных полёвок (M. gregalis) отлавливали методом ловушко-линий. У 213 полёвок исследовали изменчивость 60 неметрических признаков трёх нижних коренных зубов и комбинаций (морфотипов) жевательной поверхности первого нижнего коренного зуба (M1) [3]. В работе использовали каталог фенов и определитель морфотипов жевательной поверхности коренных зубов (рис. 2) [4; 7]. Фены и морфотипы подсчитывали на левой и правой сторонах черепа. Признаки, проявившие существенную (R≥0,3) достоверную связь с полом, возрастом, годом и местом сбора исключали из анализа [5]. Частоты фенов и морфотипов, фенетические дистанции (MMD) между выборками и их средние стандартные отклонения (MSD) рассчитывали на основе общепринятых формул [5].
Для оценки стабильности развития популяций клопа-солдатика рассчитывали показатели флуктуирующей асимметрии фенов рисунка надкрылий, для узкочерепной полёвки морфотипов M1(ФА) [6]. Описанные расчёты производили с помощью пакетов прикладных программ Statan и Phen.
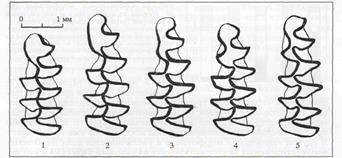
Рис. 2. Основные морфотипы первого нижнего коренного зуба узкочерепной полёвки (по А.В. Бородину, 2009)
Результаты исследования и их обсуждение
Анализ рисунка надкрылий клопа-солдатика показал более высокую изменчивость по всем элементам в городской популяции в районе мясокомбината (55 фенов), чем в популяции лесопарка (31фен) (табл. 1).
В популяции города высокий уровень вариативности характерен для элемента А, и элемента Д. Частотный состав фенов в каждой из популяций различен. Из 19 фенов по чёрному верхнему пятну (элемент А) в популяции города встречается 18, в популяции парка – 7. По чёрному нижнему пятну (элемент Д) из 21 фена, в популяции города встречается 16, а в парке –7.
В каждой из популяций выявлены господствующие фены, достоверно (при Р≤0,05) отличающиеся по встречаемости от других. Так, в популяции города преобладают фены А2, А13, В1, В10, Д4, Т1, Т2, Т6. В парке – фены А1, В1, Д1, Д4, Т1. Сходство наблюдается только по одному фену – В1.
Таблица 1. Фенетическая характеристика популяций клопа-солдатика
|
А |
Город |
Парк |
В |
Город |
Парк |
D |
Город |
Парк |
Т |
Город |
Парк |
|
А1 |
5 |
17 |
В1 |
14 |
32 |
Д 1 |
7 |
21 |
Т1 |
11 |
15 |
|
А2 |
9 |
0 |
В2 |
4 |
1 |
Д 2 |
1 |
1 |
Т2 |
9 |
5 |
|
А3 |
4 |
1 |
В3 |
1 |
1 |
Д 3 |
1 |
1 |
Т3 |
1 |
9 |
|
А4 |
1 |
10 |
В4 |
11 |
6 |
Д 4 |
17 |
22 |
Т4 |
4 |
3 |
|
А5 |
1 |
13 |
В5 |
1 |
7 |
Д 5 |
0 |
1 |
Т5 |
6 |
5 |
|
А6 |
5 |
0 |
В6 |
0 |
1 |
Д 6 |
1 |
0 |
Т6 |
9 |
5 |
|
А7 |
1 |
4 |
В7 |
0 |
2 |
Д 7 |
10 |
0 |
Т7 |
0 |
1 |
|
А8 |
1 |
0 |
В8 |
2 |
- |
Д 8 |
0 |
3 |
Т8 |
1 |
5 |
|
А9 |
0 |
3 |
В9 |
0 |
- |
Д 9 |
0 |
0 |
Т9 |
0 |
1 |
|
А10 |
2 |
1 |
В10 |
16 |
- |
Д 10 |
1 |
1 |
Т10 |
2 |
0 |
|
А11 |
3 |
- |
В11 |
0 |
- |
Д 11 |
1 |
0 |
Т11 |
4 |
0 |
|
А12 |
1 |
- |
В12 |
0 |
- |
Д 12 |
3 |
0 |
Т12 |
1 |
1 |
|
А13 |
9 |
- |
В13 |
0 |
- |
Д 13 |
0 |
- |
Т13 |
0 |
- |
|
А14 |
1 |
- |
В14 |
0 |
- |
Д 14 |
1 |
- |
Т14 |
0 |
- |
|
А15 |
0 |
- |
В15 |
1 |
- |
Д 15 |
1 |
- |
Т15 |
1 |
- |
|
А16 |
1 |
- |
|
|
|
Д 16 |
2 |
- |
Т16 |
0 |
- |
|
А17 |
1 |
- |
|
|
|
Д 17 |
0 |
- |
Т17 |
1 |
- |
|
А18 |
3 |
- |
|
|
|
Д 18 |
1 |
- |
|
|
|
|
А19 |
1 |
- |
|
|
|
Д 19 |
1 |
- |
|
|
|
|
А20 |
1 |
- |
|
|
|
Д 20 |
1 |
- |
|
|
|
|
|
- |
- |
|
|
|
Д 21 |
2 |
- |
|
|
|
|
итого |
18 |
7 |
|
8 |
7 |
|
16 |
7 |
|
12 |
10 |
Выявлены фены-маркеры. По элементу А в популяции города маркеры -А2, А6, А8, по элементу Д наблюдается 4 фена-маркера (Д6, Д7, Д11, Д12). По элементу Т маркерами являются Т10 и Т11. В парке также встречаются фены – маркеры: А9, В6, Д5, Д8, Т7, Т9. По элементу В маркеров не выявлено.
В городской популяции клопа-солдатика по 24 фенам выявлен высокий уровень направленной асимметрии, по 12 фенам – высокий уровень флуктуирующей асимметрии (табл. 2). Больше асимметричных фенов наблюдается по элементам А (6 фенов) и Д (5 фенов). Полученные результаты свидетельствуют о снижении стабильности развития в популяции города, которая испытывает интенсивную антропогенную нагрузку.
В исследуемых популяционных группах узкочерепной полёвки выявлено 60 фенов жевательной поверхности нижних коренных зубов. Каждая популяционная группа имеет специфическую фенетическую структуру, что обусловлено разным качественным составом и различной встречаемостью анализируемых фенов. Городская выборка характеризуется наиболее разнообразным составом фенов (60 признаков). В выборке из естественного местообитания отмечен 51 фен. Однообразие местообитаний на территории г. Ишима, в которых отлавливались полёвки, позволяет предположить, что разнообразие фенетической структуры ишимской популяционной группы поддерживается за счёт дестабилизирующего влияния на процессы развития животных изменённой городской среды, насыщенной загрязнителями различного происхождения.
Таблица 2. Статистические показатели стабильности развития в анализируемых популяциях
|
Город |
Парк |
||||||
|
фены |
χ2 |
Доля направленной асимметрии % |
Доля флуктуирующей асимметрии % |
фены |
χ2 |
Доля направленной асимметрии % |
Доля флуктуирующей асимметрии % |
|
А1 |
9,9 |
0 |
100 |
А1 |
20,4 |
2 |
98 |
|
А2 |
24,1 |
25 |
75 |
А2 |
20,4 |
100 |
0 |
|
А3 |
29,0 |
100 |
0 |
А3 |
20,4 |
100 |
0 |
|
А4 |
29,0 |
100 |
0 |
А4 |
17,8 |
11 |
89 |
|
А5 |
29,0 |
100 |
0 |
А5 |
30,9 |
100 |
0 |
|
А6 |
19,1 |
100 |
0 |
А7 |
10,5 |
25 |
75 |
|
А10 |
7,7 |
11 |
89 |
А8 |
10,5 |
100 |
0 |
|
А13 |
9,4 |
2 |
98 |
А9 |
17,6 |
100 |
0 |
|
А18 |
7,6 |
11 |
89 |
А10 |
5,6 |
100 |
0 |
|
А19 |
4,1 |
44 |
56 |
|
|
|
|
|
А20 |
4,1 |
100 |
0 |
|
|
|
|
|
В1 |
40,5 |
100 |
0 |
В1 |
23,6 |
51 |
49 |
|
В4 |
10,9 |
100 |
0 |
В2 |
4,9 |
44 |
56 |
|
В10 |
3,9 |
100 |
0 |
В3 |
5,6 |
100 |
0 |
|
D1 |
34,2 |
100 |
0 |
D1 |
17,2 |
1 |
99 |
|
D2 |
34,2 |
100 |
0 |
D2 |
5,6 |
100 |
0 |
|
D3 |
7,7 |
11 |
89 |
D3 |
14,1 |
0 |
100 |
|
D4 |
20,1 |
51 |
49 |
D4 |
32,3 |
100 |
0 |
|
D6 |
5,6 |
100 |
0 |
|
|
|
|
|
D7 |
5,5 |
7 |
93 |
|
|
|
|
|
D8 |
5,5 |
100 |
0 |
|
|
|
|
|
D10 |
4,7 |
36 |
64 |
|
|
|
|
|
D11 |
14,1 |
0 |
100 |
|
|
|
|
|
Т1 |
33,6 |
100 |
0 |
Т1 |
22,2 |
0 |
100 |
|
Т2 |
29,0 |
100 |
0 |
Т2 |
22,2 |
100 |
0 |
|
Т4 |
12,7 |
100 |
0 |
Т3 |
43,5 |
11 |
89 |
|
Т17 |
14,1 |
0 |
100 |
Т5 |
19,5 |
100 |
0 |
|
|
|
|
|
Т6 |
9,9 |
0 |
100 |
|
|
|
|
|
Т8 |
29,0 |
100 |
0 |
Примечание: в таблице представлены фены, по которым асимметрия подтверждена статистически.
Количественные различия между контрольной и импактной группами проявляются в различных частотах одинаковых фенов. Так, в контрольной выборке 100 %-ной встречаемостью характеризуются фены Cins1 и Sics1; на 2-м месте по встречаемости находится фен Ppt2 (92 %); на 3-м месте – фен Ali1 (76 %); высокой частотой отличатся фены Lipt1 (72 %) и Ili2 (50 %); наиболее редко (1 %) встречаются фены Pat3, Ppt3, Ain4, Phc4, Cens3, Lipt3, Ili4, Ili5. B городской выборке самым распространённым является фен Cins1 (93 %), на втором месте – фен Ali1 (79 %), на 3-м – Lipt1(75 %); чрезвычайно редки фены Fli9 (0,04 %), Ppt4 (0,04 %), очень редки – Ali7 (0,4 %), Ili7 (0,4%), редки фены Ali6 (1 %), Fli7 (1 %), Fli8 (1 %). Множественное сравнение по отдельным признакам выявило статистически значимые межгрупповые различия по 26 фенам (табл. 3).
Уровень фенетической дистанции между выборками выше, нежели определяет степень пространственно-географической изолированности [7]. Это позволяет предположить, что полученные результаты обусловлены влиянием антропогенных загрязнителей.
В изучаемых выборках узкочерепной полёвки выявлены все 5 морфотипов M1, описанные другими авторами [3-4]. Среди выявленных морфотипов обнаружены как эволюционно древние, простые без выраженного входящего угла на внешней (буккальной) стороне зуба (морфотипы I–III), так и более эволюционно молодые и сложные морфотипы (IV–V) c едва заметным или хорошо выраженным входящим углом (рис. 2).
В обеих (контрольной и опытной) выборках наибольшей встречаемостью характеризуется IV морфотип с выраженным острым углом на буккальнойстороне передней непарной петли (рис.2). Его частота составляет соответственно 0,42±0,05 и 0,38±0,05. Второе место по встречаемости в контрольной выборке занимают II и III морфотипы (их частоты равны и составляют 0,17±0,04), в городской выборке – II морфотип (0,17±0,04). На третьем месте в контрольной выборке находится I морфотип (0,16±0,04), а в городской – III морфотип (0,17±0,04). Самым редким в обеих выборках является наиболеесложный V морфотип. Его частота составляет соответственно 0,07±0,03 и 0,03±0,02. Из представленных данных видно, что выборки узкочерепной полёвки из естественного местообитания и городских лесопарков имеют сходную морфотипическую структуру и значимо различаются только по частоте встречаемости II морфотипа (t=2,35; при Р ≤0,05).
Расчёт коэффициента ранговой корреляции Спирмена показал отсутствие значимой существенной корреляции между встречаемостью различных морфотипов и локальной принадлежностью животных. Всё это служит косвенным, но достаточно определённым подтверждением адаптивного характера анализируемых морфотипов (представляющих собой дискретности второго порядка – комбинации фенов) в пределах одной физико-географической зоны.
Таблица 3. Множественное фенетическое сравнение разных популяционных групп узкочерепной полёвки
|
№ п/п |
Фены |
естественный биотоп |
городской лесопарк |
фенетическая дистанция |
χ2 |
|
1 |
Ali 2 |
0,02±0,02 |
0,33±0,03 |
-0,319 |
20,37*** |
|
2 |
Ali 4 |
0,04±0,02 |
0,13±0,02 |
-0,095 |
6,82*** |
|
3 |
Fli 1 |
0,50±0,05 |
0,19±0,03 |
0,432 |
28,3*** |
|
4 |
Fli 2 |
0,31±0,05 |
0,43±0,03 |
0,048 |
4,00* |
|
5 |
Fli 3 |
0,09±0,03 |
0,23±0,03 |
0,132 |
9,32** |
|
6 |
Cr 1 |
0,29±0,05 |
0,12±0,02 |
0,158 |
11,0*** |
|
7 |
Cr 2 |
0,71±0,05 |
0,37±0,03 |
0,471 |
30,72*** |
|
8 |
Cin 2 |
0,61±0,05 |
0,48±0,03 |
0,060 |
4,77* |
|
9 |
Pat 3 |
0,01±0,01 |
0,27±0,03 |
0,701 |
45,1*** |
|
10 |
Cem 3 |
0,09±0,03 |
0,21±0,03 |
0,093 |
6,81** |
|
11 |
Ppt 2 |
0,92±0,03 |
0,50±0,02 |
0,928 |
58,6*** |
|
12 |
Ppt 3 |
0,01±0,01 |
0,45±0,03 |
1,440 |
90,1*** |
|
13 |
Ain 1 |
0,36±0,05 |
0,22±0,03 |
0,083 |
6,12* |
|
14 |
Ain 4 |
0,01±0,01 |
0,16±0,02 |
0,305 |
19,9*** |
|
15 |
Phc 4 |
0,01±0,01 |
0,15±0,02 |
0,284 |
18,9*** |
|
16 |
Pats 1 |
0,14±0,08 |
0,05±0,01 |
0,079 |
5,96* |
|
17 |
Pats 2 |
0,43±0,05 |
0,20±0,03 |
0,234 |
15,7*** |
|
18 |
Pats 3 |
0±0 |
0,18±0,03 |
0,585 |
37,8*** |
|
19 |
Pats 4 |
0,43±0,05 |
0,29±0,03 |
0,072 |
5,54* |
|
20 |
Lips 3 |
0±0 |
0,13±0,02 |
0,387 |
25,5*** |
|
21 |
Ppts 2 |
0,09±0,03 |
0,06±0,02 |
0,859 |
54,8*** |
|
22 |
Cins 1 |
1,0±0,01 |
0,93±0,02 |
5,395 |
335,3*** |
|
23 |
Cins 2 |
0±0 |
0,48±0,03 |
1,979 |
124,3*** |
|
24 |
Sics 1 |
1,0±0 |
0,49±0,03 |
1,647 |
104,2*** |
|
25 |
Sics 2 |
0±0 |
0,50±0,03 |
2,082 |
130,7*** |
|
26 |
Cint 1 |
0,40±0,05 |
0,26±0,03 |
0,069 |
5,26* |
|
MMD |
0,308 |
χ2=1230,4*** |
|||
|
MSD |
0,0029 |
|
|||
Примечание: в таблице представлены фены, по которым выявлены достоверные межгрупповые различия; MMD – средняя фенетическая дистанция, рассчитанная по комплексу фенов; MSD – среднее стандартное отклонение.
В городской выборке M. gregalis отмечается возрастание доли флуктуирующей асимметрии по II морфотипу (c 0,02 % до 96 %). Очевидно, можно сделать вывод о дестабилизирующем влиянии городской среды на популяционный гомеорез узкочерепной полёвки.
Выводы
Исследование эпигенетической изменчивости в популяциях клопа-солдатика и узкочерепной полёвки показало, что в градиенте урбанизации возрастает эпигенетический полиморфизм.
Так, у клопа-солдатика в городской популяции, испытывающей большую антропогенную нагрузку, происходит изменение соотношения фенов, увеличивается их количество и возрастает их разнообразие, появляются фены-маркеры, которые могут служить индикаторами антропогенных загрязнителей. По элементам А и Д наблюдается повышение доли флуктуирующей асимметрии в городской популяции, что свидетельствует о снижении стабильности развития особей, обусловленной действием средового стресса.
В городской популяции узкочерепной полёвки увеличивается разнообразие фенов неметрических признаков жевательной поверхности нижних коренных зубов, изменяется встречаемость фенов. В условиях антропогенного стресса усиливается феногенетическая дифференциация популяций и возрастает флуктуирующая асимметрия по отдельным признакам, что служит показателем дестабилизации популяционного гомеореза.
Рецензенты:
Лихачёв С.Ф., д.б.н., профессор, декан факультета экологии Челябинского государственного университета, г. Челябинск.
Гашев С.Н., д.б.н., профессор, заведующий кафедрой зоологии и эволюционной экологии животных Тюменского государственного университета, г. Тюмень.
Библиографическая ссылка
Левых А.Ю., Пузынина Г.Г. ЭПИГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ НЕМЕТРИЧЕСКИХ ПРИЗНАКОВ У ЖИВОТНЫХ ИЗ ПОПУЛЯЦИЙ С РАЗНОЙ СТЕПЕНЬЮ АНТРОПОГЕННОЙ НАГРУЗКИ // Современные проблемы науки и образования. 2013. № 6. ;URL: https://science-education.ru/en/article/view?id=11441 (дата обращения: 01.07.2026).