


Scientific journal
Modern problems of science and education
ISSN 2070-7428
"Перечень" ВАК
ИФ РИНЦ = 0,936
RESEARCH OF STRESSFUL REACTIONS ЭПИДЕРМИСА OF THE LEAF OF TYPES OF SORT JUGLANS GROWING IN CONDITIONS OF THE BELGOROD AREA ON ACTION OF HEATS

Введение
Засуха, приводящая к возникновению водного дефицита в растении, вызывающая временное завядание, отражается на продуктивности растений. Особую чувствительность к засухе растения проявляют в критические периоды потребности в воде.
Для увеличения разнообразия пищевой продукции, особенно в годы с аномальными погодными условиями, проводится интродукция растений в Белгородскую область. Виды из рода Juglans наиболее интересны, так как обладают высокими вкусовыми, пищевыми и лечебными качествами [2, 3, 5]. В Белгородской области нет ни одной орехоплодной плантации, грецкий орех встречается в основном в частных хозяйствах и изредка используется в озеленении улиц.
Задача исследования – изучить роль устьиц и основных клеток адаксиального и абаксиального эпидермиса в регуляции водного обмена листа видов рода Juglans.
Материалы и методы. Объектами исследования стали растения семи видов рода Juglans, произрастающих в Ботаническом саду БелГУ, 2001 года посадки: J. regia L., J. manshurica Max., J. siboldiana Maxim., J. cordiformis Max., J. nigra L., J. cinerea L., J. rupestris Engelm. в фенофазу формирования плодов. Контроль – растения вида J. regia L., произрастающий в течение длительного времени в Белгородской области. Учитывалось происхождение видов.
Исследовались листья после воздействия температур 27, 30, 35, 40º С в течение 40 минут при влажности воздуха 54% при исключении подачи воды корневой системы. Находили интенсивность транспирации, взвешивая лист до и после термического воздействия.
Комплексная оценка засухоустойчивости осуществлялась, согласно методике Никитского ботанического сада (Кормилицын, Голубева, 1970).
Для исследования отбирались листья с годичных вегетативно-плагиотропных приростов (7-й от основания прироста) из средней части кроны, с учетом их морфологического адреса, освещенности в утренние часы. Консервация листьев проводилась общепринятым способом в смеси: спирт, вода, глицерин (1:1:1) [1]. Приготовление препаратов эпидермиса проводилось по модифицированной методике [5].
Изучение эпидермиса и его структур проводилось с помощью СМ «Биолам С 13», Микромед-5, бинокуляра МБС 10 и РЭМ Quanta 200 3D в центре коллективного пользования научным оборудованием БелГУ, программы «ВидиоТест-Мастер».
При анатомическом исследовании учитывались признаки зрелых, полностью сформированных структур при 50-кратной повторности измерений. В средней части листа между жилками подсчитывали концентрацию устьиц абаксиального эпидермиса, измеряли их длину и ширину, степень открытости устьиц (СОУ) [3], их ориентацию и степень погруженности. Находили площадь, удлиненность и степень извилистости антиклинальных стенок основных клеток адаксиального и абаксиального эпидермиса. Для этого очерченный периметр клетки делили на ее площадь. РЭМ позволил описать форму основных клеток эпидермиса в трехмерной системе координат.
При анализе полученных данных использовалась статистическая обработка по Г.Н. Зайцеву [2, 3, 5] при помощи пакета программ Microsoft Office.
Обсуждение результатов
Лист — это один из основных органов растения, выполняющий функции фотосинтеза, транспирации (испарения воды растением) и газообмена с окружающей средой. Значительную долю устойчивости растений к засухе определяет их способность противостоять температурному стрессу и водному дефициту. При высокотемпературном стрессовом воздействии наблюдаются ответные защитные реакции, позволяющие выявить наиболее стресстолерантные виды растений и определить механизмы их адаптации к стрессовому воздействию.
Термическое воздействие на листья видов рода Орех приводят к увеличению интенсивности транспирации. Наименьшая водоудерживающая способность листа при всех температурах отмечается у видов J. manshurica, J. cjhdiformis, J. siboldiana (ткани листа теряют до 30% воды от своей массы), а наименьшая – для J. regia, J. nigra (ткани листа теряют до 14% воды от своей массы).
Для видов J. manshurica, J. cjhdiformis, J. rupestris интенсивность транспирации максимальна при воздействии 40С, а при воздействии 45С потеря влаги существенно снижается. Для J. siboldiana, J. regia, J. nigra, J. cinerea интенсивность транспирации наибольшая при воздействии 45С.
Водоудерживающая способность имеет прямую зависимость от соотношения свободной и связаной воды (рис. 1).
А 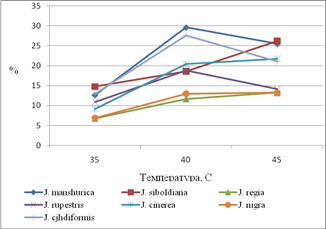 В
В 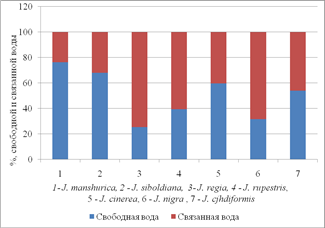
Рис. 1 Водоудерживающая способность в зависимости от соотношения свободной и связаной воды.
По нашим данным, J. manshurica, J. cоrdiformis, J. siboldiana, J. cinerea имеют наибольшую долю свободной воды по отношению к оводненности тканей листа, чем у других видов. Наиболее информативным ксероморфным признаком у видов рода Juglans являются особенности распределения кутикулярного слоя по поверхности абаксиального и адаксиального эпидермиса, степень открытости устьиц и их погруженность. Большое значение имеет утолщение кутикулярного слоя между выростами основных клеток эпидермиса, а также особенность распределения кутикулы вокруг устьиц. Устьица на абаксиальной поверхности J. regia, J. nigra, J. rupestris отличаются утолщенным на поверхности слоем кутикулы, устьица J. manshurica, J. cоrdiformis, J. siboldiana, J. cinerea находятся на поверхности эпидермиса и не имеют выраженного кутикулярного слоя.
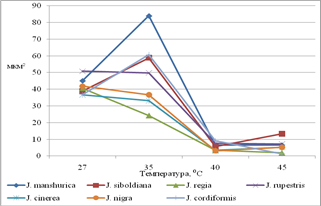
Реакция устьиц на изменение температуры при постоянной влажности воздуха (54%). Состояние устьичного аппарата в значительной степени определяет водный баланс растений и интенсивность ассимиляции СО2, что отражается на характере продукционного процесса. При нарастании засухи наблюдается снижение устьичной проводимости. В условиях глубокой засухи возможны нарушения структуры устьичного аппарата, и снижалась способность устьиц регулировать ширину устьичной щели. При нарастании засухи наблюдается изменение интенсивности фотосинтеза, что обусловлено закрыванием устьиц и уменьшением поглощения СО2, что ведет к падению синтеза крахмала и сахаров. Высокая интенсивность транспирации обусловлена увеличением устьичной проводимости. При 27С наибольшая устьичная проводимость наблюдалась у о. Зибольда, о. маньчжурского, о. сердцевидного, при 35-40С наибольшая СОУ у о. маньчжурского и сердцевидного, что сопоставимо с высокой интенсивностью транспирации листьев этих видов при воздействии температур (рис. 2), а также наибольшей долей свободной воды в тканях листа (рис. 1). Среди изучаемых видов наименьшей интенсивностью транспирации и пропускной способностью устьиц обладают листья ореха грецкого (рис. 2). Доля свободной воды в тканях листа среди изучаемых видов наименьшая у ореха грецкого (рис. 1).
Для грецкого ореха потеря воды незначительна, не превышает 12 % от массы листа. Проводимость устьиц снижается при повышении температуры на 8С. Держится на одном уровне при 35-40С и значительно уменьшается при 45С. Устьица полностью закрываются при уже 40С. Площадь замыкающих клеток увеличивается к 35С и резко падает к 40С. Таким образом, устьица листьев о. грецкого обладают высокой термореактивностью и некоторой устойчивостью тургора замыкающих клеток при температуре 27-35С.
Орех черный характеризуется также высокой реактивностью устьиц и поддержанием тургора замыкающих клеток при действии температур 27-35С. Однако при критическом термическом воздействии (45С) нарушается регуляторная способность устьиц, они открываются.
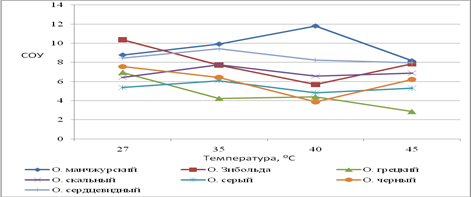
Рис. 2. Степень открытости устьиц
Орех скальный характеризуется незначительным уменьшением проводимости устьиц при действии температур 27-45С. Потеря влаги не превышает 20%, снижается при действии 45С. С повышением силы стрессового воздействия уменьшается размер замыкающих клеток (вследствие снижения тургора), а также закрывается устьичная щель. Таким образом, при 27-35С проводимость устьиц не уменьшается, сохраняются условия для оптимального газообмен и фотосинтеза. Ткани о. Зибольда при увеличении стрессового воздействия температур увеличивают потерю воды (до 30% от массы листа). При повышении температуры на 8°С площадь устьичной щели и замыкающих клеток растет при действии 27-35° С и резко снижается при 40-45С. Ткани Ореха серого при повышении температуры увеличивают потерю воды (до 22% от массы листа). СОУ практически не изменяется при увеличении стресса. Размеры замыкающих клеток и устьичной щели значительно снижаются к 40°С. Орех маньчжурский отличается высокой потерей влаги при действии температур (27-45°С). Наибольшая потеря влаги при действии 40°С (30% от массы листа). СОУ ореха маньчжурского достоверно больше, чем у о. грецкого. Проводимость устьиц увеличивается к 40°С, только при действии 45°С снижается. При 35С пропускная способность устьиц увеличивается (растет площадь замыкающих клеток и устьичной щели). При 40-45°С площадь устьица и устьичной щели значительно снижается (рис. 3).
Ткани листа ореха сердцевидного при температурном воздействии теряют до 30% влаги от массы листа. СОУ при температурном воздействии незначительно снижается. Площадь устьичной щели и замыкающих клеток растет при действии температур до 35° С и значительно снижается при действии температур 40-45°С.
В нашем исследовании не всегда наблюдается отрицательная корреляция между нарастанием водного дефицита и проводимостью устьиц. У о. Зибольда и черного с увеличением водного дефицита устьица не снижают свою проводимость. По литературным данным, в условиях глубокой засухи нарушается структура устьичного аппарата и снижается способность устьиц регулировать ширину устьичной щели.
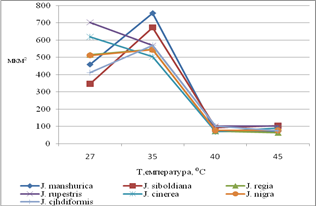
Площадь замыкающих клеток
Площадь устьичной щели
Рис. 3. Некоторые особенности морфологии листа.
Реакция основных клеток адаксиального и абаксиального эпидермиса на изменение температуры при постоянной влажности воздуха (54%).
Основное внимание исследователей сосредоточено на изучении роли устьиц в регуляции водного обмена [2, 3] . При температурном воздействии мы исследовали изменения, происходящие с основными клетками абаксиального и адаксиального эпидермиса при потере тканями листа влаги.
Уменьшение тургора при потере влаги ведет к уменьшению площади основных клеток, этому процессу препятствует клеточная стенка, а также кутикулярный слой на поверхности эпидермиса. Самые значительные уменьшения площади клеток происходит при действии температур 40-45С.
Основные клетки адаксиального эпидермиса J. manshurica при увеличении стрессового воздействия значительно снижают свою площадь уже при действии 35С, что свидетельствует о более низкой устойчивости к влагопотере. Причиной такой реакции может быть отсутствие кутикулярного слоя на поверхности эпидермиса, а также тонкая клеточная стенка. Площадь основных клеток адаксиального эпидермиса J. siboldiana, J. nigra, J. cordiformis, J. rupestris при действии 35°С несколько увеличивается. Эти клетки имеют выпуклую проекцию, при снижении тургора клетки, уплощаясь, увеличивают тангентальную поверхность.
Площадь основных клеток абаксиального эпидермиса при действии 35°С незначительно увеличивается и существенно уменьшается при 40-45°С. Больше всех варьирует площадь основных клеток абаксиального эпидермиса J. manshurica, что объясняется отсутствием кутикулярного слоя на поверхности эпидермиса и тонкой клеточной стенкой.
Снижение тургорного давления в клетках ведет к увеличению извилистости антиклинальных стенок. Наиболее значимые изменения коэффициента извилистости антиклинальных стенок наблюдаются у основных клеток J. manshurica, что подтверждает предположение об уменьшении сопротивления клеточной стенки большой влагопотере, а эти признаки – показатель действия засухи, высоких температур, водного дефицита, а признаком ксероморфности является способность сохранять высокую продуктивность в засушливых условиях.
Выводы
1. Наименьшей водоудерживающей способностью обладают листья J. manshurica, J. siboldiana, J. cordiformis, что сопоставимо с большой долей свободной воды в тканях листа; наибольшей водоудерживающей способностью обладают листья J. regia, J. nigra.
2. Устьица J. regia J. nigra, J. cinerea обладают высокой термореактивностью, проводимость устьиц значительно снижается к 40С, устьица имеют выраженный кутикулярный слой на поверхности. Устьица J. manshurica, J. siboldiana, J. cordiformis характеризуются большой проводимости при действии 35С, отличительной особенность устьиц – отсутствие выраженного кутикулярного слоя на поверхности, выступающее их положение над поверхностью эпидермиса.
3. Основные клетки абаксиального и адаксиального эпидермиса при потере влаги уменьшают площадь, увеличивается коэффициент извилистости антиклинальных стенок. Увеличение толщены клеточной стенки, выраженности кутикулярного слоя на поверхности основных клеток увеличивает устойчивость клеток эпидермиса к влагопотере. Основные клетки J. manshurica отличаются наименьшей устойчивостью к водному дефициту (особенной основные клетки адаксиального эпидермиса).
Рецензенты:
Ткаченко И.К., д.с.-х.н., профессор, профессор кафедры анатомии и физиологии живых организмов Биолого-химического факультета Белгородского государственного университета Министерства образования и науки РФ, г. Белгород.
Лазарев А.В., д.б.н., доцент, профессор кафедры биотехнологии и микробиологии Биолого-химического факультета Белгородского государственного университета Министерства образования и науки РФ, г. Белгород.
Библиографическая ссылка
Назарова Н.В., Кузнецова Т.А., Сорокопудов В.Н., Шестопалова Н.Н. ИССЛЕДОВАНИЕ СТРЕССОВЫХ РЕАКЦИЙ ЭПИДЕРМИСА ЛИСТА ВИДОВ РОДА JUGLANS, ПРОИЗРАСТАЮЩИХ В УСЛОВИЯХ БЕЛГОРОДСКОЙ ОБЛАСТИ, НА ДЕЙСТВИЕ ВЫСОКИХ ТЕМПЕРАТУР // Современные проблемы науки и образования. 2013. № 6. ;URL: https://science-education.ru/en/article/view?id=11131 (дата обращения: 17.07.2026).