



Рак яичников является ведущей причиной летальных исходов от гинекологических злокачественных опухолей в мире и обычно диагностируется уже на поздней стадии. Серозная аденокарцинома является одним из самых распространенных подтипов эпителиальных опухолей яичников [1]. Число летальных исходов от этой патологии является самым высоким среди гинекологических опухолей в мире и ежегодно увеличивается. В исследовании Blagden S.P. (2015) установлено, что серозная аденокарцинома яичника гетерогенна на гистологическом и молекулярном уровнях [2]. Традиционно выделяли два подтипа этого рака: серозную аденокарциному высокой степени злокачественности (high-grade serous cancer) и серозную аденокарциному низкой степени злокачественности (low-grade serous cancer) [3]. Для серозной аденокарциномы яичника низкой степени злокачественности свойственно относительно благоприятное клиническое течение, а также стабильный молекулярно-генетический и эпигенетический профиль. Серозная аденокарцинома яичника высокой степени злокачественности имеет агрессивное клиническое течение [1], при этом для неё в настоящее время не существует одобренного скринингового теста, что препятствует ранней диагностике данного заболевания [4].
Для разработки эффективных и малоинвазивных методов ранней диагностики необходим скрининг молекулярных маркеров во внеклеточной ДНК плазмы крови. В качестве таких маркеров большим потенциалом обладает показатель числа копий генов (Copy Number Variation (CNV)) - полиморфизм, приводящий к изменению копийности генетического локуса и, как следствие, изменению экспрессии этого локуса и его продукта - белка или не кодирующей РНК [5]. Показатель CNV может выступать в качестве высокоспецифичного биологического маркера, позволяющего осуществлять как раннюю диагностику [5], так и молекулярное типирование опухолей [6].
К внеклеточной ДНК (внДНК) относится ядерная и митохондриальная ДНК из соматических и опухолевых клеток, подвергшихся апоптозу или некрозу; ДНК из клеток крови, вирусная и бактериальная ДНК [7]. Проведенный анализ баз данных TCGA и DisGeNET позволил выделить ряд генов (регулирующих метаболизм эстрогена), показатель копийности которых имеет потенциал малоинвазивной диагностики серозной аденокарциномы яичников (табл. 1).
Таблица 1
Перечень потенциальных молекулярных маркеров серозной аденокарциномы яичников
|
№ |
Название гена |
Варианты названия гена |
Расшифровка |
|
1 |
CYP1-A1 |
CYP1, P450DX, CP11, P1-450, AHRR, AHH, CYPIA1, P450C |
цитохром P450 семейство 1 подсемейство A1 |
|
2 |
CYP1-A2 |
CYPIA2, P450(PA), CP-12, P3-450 |
цитохром P450 семейство 1 подсемейство A2 |
|
3 |
CYP1-B1 |
CYPIB1, P4501B1, ASGD6, GLC3A, CP1B |
цитохром P450 семейство 1 подсемейство B1 |
|
4 |
CYP19A1 |
ARO1, CPV1, CYP19, ARO, P-450AROM, CYAR |
цитохром P450 семейство 19 подсемейство A1 |
|
5 |
ESR1 |
ESR, NR3A1, Era, ESTRR, ER, ESRA |
рецептор эстрогена 1 |
|
6 |
ESR2 |
NR3A2, Erb, ER-BETA, ESRB, ESTRB, |
рецептор эстрогена 1 |
|
7 |
GPER |
LERGU, CEPR, FEG-1, LyGPR, LERGU2, GPCR-Br, CMKRL2, GPER, mER, DRY12, GPR30 |
рецептор эстрогена 1, связанный с G-белком |
|
8 |
STS |
ARSC1, ASC, ARSC, XLI, SSDD, ES |
стероидная сульфатаза |
|
9 |
SULT1-A1 |
P-PST, ST1-A1, PST, STP, TSPS-T1, HAST1/2, STP-1, ST1-A3 |
семейство сульфотрансфераз 1 подсемейство A1 |
|
10 |
SULT1-E1 |
EST-1, ST1-E1,EST, STE |
семейство сульфотрансфераз 1 подсемейство E1 |
Целью данного исследования стало определение во внДНК показателя копийности генов, регулирующих метаболизм и рецепцию эстрогенов, у больных серозной аденокарциномой яичников высокой степени злокачественности и у условно здоровых доноров.
Материалы и методы исследования
В работе использовали кровь, взятую путем венопункции у 50 больных серозной аденокарциномой яичников высокой степени злокачественности, а также у 30 доноров без онкологической патологии (группа условно здоровых доноров).
Плазму крови отделяли центрифугированием (30 минут, 2000 об/мин, t=10 °С). Из плазмы выделяли внДНК фенол-хлороформным методом в модификации Кутилина Д.С. и соавторов [7]: к 4 мл плазмы добавляли 4 мл лизирующего раствора, содержащего 2% SDS, 1% меркаптоэтанол и протеиназу К, инкубировали 30 мин. при 58 °С, после чего добавляли 4 мл смеси щелочной фенола с хлороформом (1:1) и центрифугировали 20 мин. при 3000 об/мин (t=10 °С). Раствор разделялся на две фазы - фенол-хлороформную и водную фазу, содержащую ДНК. Водную фазу переносили в отдельную пробирку и добавляли равный объем изопропилового спирта (96%) и хлорида натрия (до концентрации в растворе 100 мМ). После 60-минутной инкубации при -20 °С проводилось центрифугирование (12 000 об/мин, 15 минут, t=10 °С). Получившийся осадок промывали этиловым спиртом (80%). Этанол удаляли центрифугированием, высушивали осадок при 58 °С и растворяли в буфере, содержащем ЭДТА [7].
Оценку показателя относительной копийности 10 генов (ESR1, GPER, ESR2, CYP1A1, CYP1A2, CYP1B1, CYP19A, SULT1A, SULT1E1, STS) проводили методом количественной ПЦР в режиме реального времени (RT-qPCR). Разработка последовательности синтетических олигонуклеотидов (праймеров) осуществлялась с помощью Primer-BLAST и базы данных GenBank (NCBI). В качестве референсного гена для нормализации полученных показателей RT-qPCR был выбран B2M.
Реакция амплификации проводилась на термоциклере CFX96 (Bio-Rad, США) в 20 мкл смеси, содержащей матрицу внДНК (не менее 0,5 нг), 0,2 мМ раствор dNTP, 0,4 мкМ прямого и обратного праймеров, 2,5 мМ раствор MgCl2, 1X ПЦР-буфер с интеркалирующим красителем EvaGreen и 0,1 е.а./мкл SynTaq ДНК-полимеразы [7]. Амплификация осуществлялась по следующей программе: 95 °C 3 минуты, 40 циклов: 95 °C 10 секунд, 55 °C 30 секунд (чтение оптического сигнала по каналу FAM) и 72 °C 20 секунд.
Относительную копийность (rC) рассчитывали по формуле, описанной Кутилиным Д.С. [8]:
rC = rCо/rCн = E-ΔCt(образцов от больных)/E-ΔCt(образцов от условно здоровых),
где Е ‒ эффективность ПЦР-амплификации, рассчитанная по формуле E = 10-1/k (k ‒ коэффициент уравнения прямой C(T) = k•log P0 + b, полученной в ходе линейной аппроксимации данных экспериментальных постановок ПЦР) (усредненное значение Е = 1,914), а ΔCt - разница между средним геометрическим Ct(гена-мишени) и средним геометрическим Ct(референсного гена) [8].
Статистическую обработку данных осуществляли с использованием Microsoft Excel 2010, Statistica 10.0 и среды для программирования R i386 4.0.0. Статистическую значимость различий оценивали по критерию Манна-Уитни, для корректировки множественного сравнения применяли поправку Бонферрони. Для выявления общих сигнальных путей исследуемых генов использовали алгоритм «сетевой интеграции нескольких ассоциаций», который предсказывает функцию и положение гена в составе сложной сети из множества других генов, а также рассчитывает оценку (W-value) для каждой точки построенной сети, отражающую силу связи между соседними точками [9].
Результаты исследования и их обсуждение
В ходе исследования обнаружено статистически значимое (р<0,005) увеличение копийности генов SULT1E1, CYP1B1 и ESR1 в 2,8 раза, 3,0 раза и 5.0 раз соответственно во внеклеточной ДНК плазмы крови у 80%, 75% и 85% соответственно обследованных больных серозной аденокарциномой яичников высокой степени злокачественности относительно внеклеточной ДНК плазмы крови условно здоровых доноров (рис. 1).
У 40% больных во внеклеточной ДНК наблюдалось увеличение копийности генов CYP1A1 и CYP1A2 в 2,4 раза (р<0,05) и 2,5 раза (р<0,05) соответственно, ещё у 40% больных отсутствовали изменения в копийности этих генов относительно условно здоровых доноров, а у 20% больных наблюдалось снижение копийности этих генов в 1,3 (р<0,05) и 1,4 раза (р<0,05) соответственно.
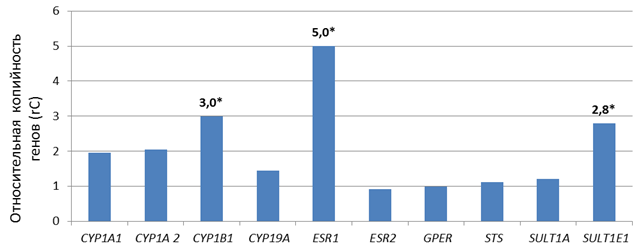
Рис. 1. Уровень относительного количества копий генов во внДНК плазмы крови больных серозной аденокарциномой яичников высокой степени злокачественности.
*– статистически значимые отличия относительно количества копий генов во внДНК плазмы крови условно здоровых доноров (р<0,005)
Копийность генов CYP19A, GPER, ESR2, STS и SULT1A статистически значимо не отличается во внеклеточной ДНК плазмы крови больных серозной аденокарциномой яичников высокой степени злокачественности и во внеклеточной ДНК плазмы крови условно здоровых доноров.
С использованием алгоритма «сетевой интеграции нескольких ассоциаций» было оценено наличие и сила взаимодействия между генами ESR1, ESR2, GPER, CYP1A1, CYP1A2, CYP1B1, CYP19A, STS, SULT1A, SULT1E1, а также генами, ассоциированными с ними в общих сигнальных путях (рис. 2, табл. 2). Для генетических локусов (SULT1A1, CYP1B1, CYP19A1, ESR2, ESR1, SULT1E1 и CYP1A1) наиболее сильное взаимодействие (определено по нескольким параметрам: совместная экспрессия генов (база Gene Expression Omnibus), общие белковые домены (базы BioGRID, IntAct и MIPS), общая регуляция транскрипционными факторами (JASPAR), со-локализация) выявлено с генами SULT1A2, CYP1A1, CYP11A1, REXO4, CYP1B1 и CYP2D6. Соответственно, повышение копийности генов ESR1, CYP1B1 и SULT1E1, помимо экспрессии соответствующих генов, может отразиться и на экспрессии генов OR52A1, ZNF398, PRDM2 и ESR2 (для ESR1), CYP1A1 и ESR1 (для CYP1B1), SULT1A1 и SULT2B1 (для SULT1E1) (рис. 2, табл. 2).
Таблица 2
Сила взаимосвязей между генами, рассчитанная с помощью алгоритма «сетевой интеграции нескольких ассоциаций»
|
Ген 1 |
Ген 2 |
W-value |
Тип взаимодействия |
|
SULT1A2 |
SULT1A1 |
0,8518850 |
предсказанные взаимодействия |
|
CYP1A1 |
CYP1B1 |
0,6785801 |
предсказанные взаимодействия |
|
CYP11A1 |
CYP19A1 |
0,4789596 |
предсказанные взаимодействия |
|
REXO4 |
ESR2 |
0,2773180 |
общие сигнальные пути |
|
CRHBP |
ESR2 |
0,2773180 |
общие сигнальные пути |
|
CRYZ |
ESR2 |
0,2773180 |
общие сигнальные пути |
|
CD68 |
ESR2 |
0,2679962 |
общие сигнальные пути |
|
CD14 |
SULT1A1 |
0,2509112 |
предсказанные взаимодействия |
|
CYP2D6 |
CYP1A1 |
0,2334536 |
предсказанные взаимодействия |
|
CYP2E1 |
CYP19A1 |
0,2142480 |
предсказанные взаимодействия |
|
CYP2D6 |
CYP1A2 |
0,2033754 |
предсказанные взаимодействия |
|
CYP1A2 |
CYP1A1 |
0,2005719 |
предсказанные взаимодействия |
|
ESR1 |
CYP1B1 |
0,1960994 |
общие сигнальные пути |
|
OR52A1 |
ESR1 |
0,1905680 |
предсказанные взаимодействия |
|
ZNF398 |
ESR1 |
0,1905680 |
предсказанные взаимодействия |
|
PRDM2 |
ESR1 |
0,1905680 |
предсказанные взаимодействия |
|
CYP3A4 |
CYP2D6 |
0,1621384 |
предсказанные взаимодействия |
|
PELP1 |
ESR2 |
0,1552160 |
предсказанные взаимодействия |
|
CYP2C18 |
CYP1A2 |
0,1467650 |
предсказанные взаимодействия |
|
CYP3A4 |
CYP1A2 |
0,1393014 |
предсказанные взаимодействия |
|
PNRC1 |
ESR2 |
0,1286611 |
общие сигнальные пути |
|
NCOA7 |
ESR2 |
0,1219798 |
предсказанные взаимодействия |
|
CYP2E1 |
CYP1A1 |
0,1155651 |
предсказанные взаимодействия |
|
CYP2C9 |
CYP2D6 |
0,1069237 |
предсказанные взаимодействия |
|
CYP1A2 |
CYP1A1 |
0,0424047 |
совместная экспрессия |
|
CYP11B2 |
CYP11A1 |
0,0343799 |
общие белковые домены |
|
ESR1 |
ESR2 |
0,0332440 |
общие белковые домены |
|
SULT1A1 |
SULT1E1 |
0,0308099 |
общие белковые домены |
|
SULT2B1 |
SULT1E1 |
0,0308099 |
общие белковые домены |
Хорошо известно, что в патогенезе ряда гинекологических злокачественных опухолей (в том числе и серозных аденокарцином яичника высокой степени злокачественности) важную роль играет эндогенная или экзогенная гиперэстрогения [1; 10]. Эстрогены образуются из андрогенов под действием ароматазы - НАДФН-зависимого фермента цитохрома Р450 19-го семейства, кодируемого геном CYP19 [11]. В данном исследовании показатель копийности гена CYP19 статистически значимо не отличается от показателя у условно здоровых доноров. Это может свидетельствовать о том, что особых изменений в процессе образования эстрогенов у больных серозной аденокарциномой яичников, вероятно, не происходит.
Эстрогены реализуют своё действие через рецепторы эстрогена, имеющиеся в разных тканях. В настоящее время известно 2 типа эстрогеновых рецепторов: α и β, функции которых интенсивно изучаются [12]. Так, в работах Bardin A. с соавторами и Кита О.И. с соавторами установлено, что гиперэкспрессия гена ESR1 сопровождает процессы онкотрансформации [13], а ESR2 регулирует митотическую активность, защищая от аномальной пролиферации индуцированной активацией α эстрогеновых рецепторов [11].
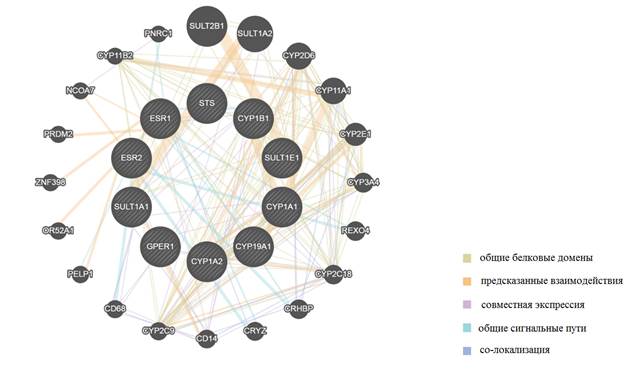
Рис. 2. Визуальная модель сети функциональных взаимосвязей между генами, регулирующими метаболизм и рецепцию эстрогенов [9]
По данным Кита О.И. и соавторов [10; 14], ещё одним ключевым фактором в канцерогенезе может быть метаболическая активация эстрадиола. Гены CYP1A1 и CYP1B1 кодируют два фермента из семейства цитохромов Р450, которые катализируют гидроксилирование 17-β-эстрадиола в положении С-2 и С-4 соответственно. Метаболиты CYP1B1 (4-гидрокси эстрогены) играют важную роль в злокачественной трансформации. Из данных литературы известно, что в некоторых гинекологических опухолях повышена экспрессия генетических локусов, регулирующих гидроксилирование эстрадиола (CYP1A1 и CYP1B1), уровень ядерных (ESR1, SULT1E1 - гены сульфотрансферазы) и мембранных рецепторов эстрогенов (GPER) [15].
Отличия копийности описанных выше генов во внДНК больных серозной аденокарциномой и относительно условно здоровых доноров вписываются в теорию эстроген-ассоциированного канцерогенеза [10], согласно которой метаболическая активация эстрадиола, вызванная аномально высокой экспрессией генов CYP1A1 и CYP1B1, и соответственно активностью кодируемых ими ферментов, в совокупности с увеличением количества ядерных рецепторов ERα, вызванным повышенной копийностью и экспрессией гена ESR1 [11], приводит к усилению пролиферации в тканях-мишенях. Этому явлению может препятствовать увеличение активности фермента сульфотрансферазы (ассоциированное с повышением экспрессии гена SULT1E1), участвующей в инактивации эстрогенов путем их сульфатирования [11]. Однако обнаруженное в нашем исследовании повышение копийности SULT1E1 не позволяет однозначно говорить о биологическом значении данного процесса (повышении экспрессии соответствующего гена и его продукта). Тем не менее показатель копийности SULT1E1 имеет потенциал в качестве молекулярного маркера серозной аденокарциномы яичника высокой степени злокачественности.
Заключение
Таким образом, проведенный анализ показателя копийности генов во внеклеточной ДНК больных серозной аденокарциномой яичников высокой степени злокачественности позволил выявить наиболее характерные малоинвазивные маркеры этого заболевания - SULT1E1, CYP1B1 и ESR1.
Библиографическая ссылка
Цандекова М.Р. ПОКАЗАТЕЛЬ КОПИЙНОСТЬ ГЕНОВ, РЕГУЛИРУЮЩИХ МЕТАБОЛИЗМ И РЕЦЕПЦИЮ ЭСТРОГЕНОВ, ВО ВНЕКЛЕТОЧНОЙ ДНК КАК ПОТЕНЦИАЛЬНЫЙ МАРКЕР ДЛЯ ДИАГНОСТИКИ СЕРОЗНОЙ АДЕНОКАРЦИНОМЫ ЯИЧНИКА ВЫСОКОЙ СТЕПЕНИ ЗЛОКАЧЕСТВЕННОСТИ // Современные проблемы науки и образования. 2020. № 5. ;URL: https://science-education.ru/ru/article/view?id=30093 (дата обращения: 19.07.2026).
DOI: https://doi.org/10.17513/spno.30093