



Во всем мире среди онкологических заболеваний рак толстой и прямой кишок занимает 4-е место по числу летальных исходов. Так, в 2018 г. было зарегистрировано более 310 000 смертей от рака прямой кишки [1].
По причине высокой распространенности разработка методов и подходов для лечения данных нозологий является одним из актуальных вопросов онкологии. Лучевая терапия (ЛТ) при колоректальном раке играет важную роль как в облегчении симптомов, так и в локальном контроле заболевания. ЛТ снижает риск возникновения локального рецидива и вызывает уменьшение размеров опухоли. Однако полного клинического ответа удается достичь в крайне малом числе случаев. Основная проблема, в которой заключается столь низкий ответ опухолей на лечение, – радиорезистентность опухолевых клеток. Хотя ряд маркеров был предложен в качестве предикторов ответа на химиолучевую терапию, ни один из них пока не используется в клинической практике. Поскольку данная группа заболеваний является гетерогенной, очень сложно найти универсальный молекулярный маркер для прогнозирования результата лечения. Эксперименты in vitro с использованием клеточных линий выявили потенциал некоторых генетических локусов (XRCC3, XRCC2, FGFR4 и NF-KB) в отношении чувствительности к ЛТ, однако все равно остается необходимость в дальнейшем изучении радиорезистентности опухолевых клеток [2, 3, 4].
Поэтому целью исследования стал скрининг предикторов радиорезистентности колоректального рака в модельном эксперименте на основании данных по аберрантной экспрессии генов, регулирующих систему сигнальных каскадов, вовлеченных в репарацию ДНК, регуляцию клеточного цикла и апоптоза.
Материалы и методы исследования
В исследовании использовалась культура клеток человека HT-29. С применением набора COrDIS Sprint проводилось генотипирование клеток для проверки соответствия стандартному генотипу данной клеточной линии и отсутствия контаминации. Культивирование клеток проводилось в стерильных флаконах (RPMI-1640, 10%-ная фетальная телячья сыворотка, гентамицин 50 мкг/мл) при 5% CO2, 95% влажности и 370C в CO2-инкубаторе CB-150 [5].
Для модельного эксперимента облучение проводили 5 раз через каждые 24 часа на линейном ускорителе Novalis TX (Varian, США) в дозах 5 и 7 Гр. Томограф Somatom Definition AS Siemens использовали на этапе топометрии [5].
Общее количество клеток, а также соотношение живых и мертвых клеток оценивали в камере Горяева, используя 0,4%-ный раствор трипанового синего. После пятого облучения клеточную линию снимали с подложки стерильного флакона раствором Трипсин/Версена. Количество клеток, находящихся на различных стадиях апоптоза, определяли на цитофлюориметре FacsCantoII (BD, USA) с использованием Annexin V-FITC Apoptosis Detection Kit [5].
Клеточную массу (культуру клеток HT-29) отмывали от среды фосфатно-солевым буферным раствором Дюльбекко (DPBS) и осаждали центрифугированием, к осадку добавляли 900 мкл реагента QIAzol (QIAGEN). Дальнейшее выделение и очистку фракции тотальной РНК производили с помощью набора RNeasy Plus Universal Kits (QIAGEN) согласно протоколу производителя. Препараты готовой РНК обрабатывали ДНКазой I для устранения следов геномной ДНК. В конечный препарат тотальной РНК добавляли РНКазин до конечной концентрации в растворе 1 ед.акт/мкл.
Качество полученной РНК проверяли электрофоретическим методом в агарозном геле по соотношению интенсивности свечения полос 28S и 18S рРНК (рис. 1).
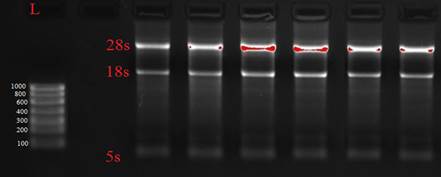
Рис. 1 Электрофореграмма РНК, выделенной из свежезамороженных тканей опухоли прямой кишки. Для визуализации использована система гель-документирования Gel Doc XR PLUS (BioRad, USA)
Концентрацию полученных препаратов РНК измеряли на флюориметре Qubit 2.0® (Invitrogen, США).
Для наработки кДНК готовили реакционную смесь, содержащую 5 мкМ рандомных праймеров, 1х RT буфер, 0,5мМ dNTP микс, 0,5 ед.акт./мкл RNase Inhibitor (Thermo scientific), 5 U/мкл ReverseTranscriptase ММLV (Синтол, Россия) и выделенную РНК в качестве матрицы. Полученную смесь инкубировали при 44°С в течение 1 часа, обратную транскриптазу инактивировали при 92°С в течение 10 минут.
Методом количественной ПЦР в режиме реального времени (RT-qPCR) определяли величины транскрипционной активности 32 генетических локусов (ATM, AKT, BRCA1, BRCA2, BRIP, CDK1, CDKN1B, CCND1, CCND3, FGFR2, HIST1, H2AX, KU70, EXO1, PTEN, RAD50, RAP80, RNF168, TOPB1, RIF1TP53, MDM2, XRCC4, BAX, BCL2, CASP8, CASP3, CASP9, RBBP8, EP300, LIG4, C-FLIP). Стабильность референсных генов оценивали с помощью алгоритма geNorm. В итоге в качестве референсных генов использовали GAPDH, ACTB и B2M. Дизайн специфичных олигонуклеотидных праймеров осуществляли с использованием NCBI GenBank и программы Primer-BLAST Последовательность праймеров представлена в таблице 1.
Таблица 1
Панель праймеров для определения относительной экспрессии генов
|
№ |
Ген |
Последовательности праймеров 5’→3’ |
|
1 |
BRCA1 |
F: ACC TGT CTC CAC AAA GTG TGA |
|
R: ACA CTG TGA AGG CCC TTT CT |
||
|
2 |
BRCA2 |
F: AGT TGG CTG ATG GTG GAT GG |
|
R: GGA TCC ACA CCT GGA GTG TC |
||
|
3 |
PTEN |
F: GGC ACA AGA GGC CCT AGA TA |
|
R: CTT AGC GCC TCT GAC TGG G |
||
|
4 |
CASP3 |
F: CTG GAA TAT CCC TGG ACA ACA TT |
|
R: TCA ACA TCT GTA CCA GAC CGA |
||
|
5 |
CASP8 |
F: CTG AAG CAA ACA GCC AGT GG |
|
R: GAT CTC AAT TCT GAT CTG CTC AC |
||
|
6 |
GAPDH |
F: GTC AAG GCT GAG AAC GGG AA |
|
R: TCG CCC CAC TTG ATT TTG GA |
||
|
7 |
BAX |
F: GGG ACG AAC TGG ACA GTA ACA |
|
R: GCT GCC ACT CGG AAA AAG AC |
||
|
8 |
B2M |
F: AGA TGA GTA TGC CTG CCG TG |
|
R: CTA TGA TGC TGC TTA CAT GTC TC |
||
|
9 |
BCL2 |
F: GGA TCC AGG ATA ACG GAG GC |
|
R: GAA ATC AAA CAG AGG CCG CA |
||
|
10 |
CASP9 |
F: TGA GAC CCT GGA CGA CAT CT |
|
R: TCC CTT TCA CCG AAA CAG CA |
||
|
11 |
P53 |
F: TTG GAA CTC AAG GAT GCC CA |
|
R: CGG GAG GTA GAC TGA CCC T |
||
|
12 |
MDM2 |
F: TAG GAG ATT TGT TTG GCG TGC |
|
R: CCT GCT GAT TGA CTA CTA CCA A |
||
|
13 |
AKT1_V1 |
F: AGCTGGTGCATCAGAGGCTG |
|
R: TGTAGCCAATGAAGGTGCCA |
||
|
14 |
ATM |
F: TGCGTGGCTAACGGAGAAAA |
|
R: ATCACTGTCACTGCACTCGG |
||
|
15 |
BRIP1 |
F: TTACCCGTCACAGCTTGCTA |
|
R: CTCATCTGCTGGTTTCCCACT |
||
|
16 |
CDK1 |
F: AAGCCGGGATCTACCATACC |
|
R: CATGGCTACCACTTGACCTGT |
||
|
17 |
CDKN1B |
F: TAATTGGGGCTCCGGCTAAC |
|
R: GAAGAATCGTCGGTTGCAGGT |
||
|
18 |
CCND1 |
F: GATCAAGTGTGACCCGGACT |
|
R: CTTGGGGTCCATGTTCTGCT |
||
|
19 |
CCND3 |
F: GTGGAGACTGGCTCTGTTCG |
|
R: TCACATACCTCCTCGTCAGGT |
||
|
20 |
FGFR2 |
F: AACAGTCATCCTGTGCCGAA |
|
R: TGGACTCAGCCGAAACTGTTA |
||
|
21 |
KU70 |
F: ACGTAGAGGGCGTTGATTGG |
|
R: TGGCTACTGCTCACTTTGGC |
||
|
22 |
RAD50 |
F: GCGTGCGGAGTTTTGGAATAG |
|
R: TTGAGCAACCTTGGGATCGT |
||
|
23 |
RAP80 |
F: GAGTGAGCAGGAAGCTAGGG |
|
R: AGAAGGCCGGCAACTATTCA |
||
|
24 |
EXO1 |
F: GAACAAGCCGGGGTTACAGA |
|
R: AGGAGGAAGCTTTTCAGAATTTTT |
||
|
25 |
Rif1 |
F: GGCTGTTTCCATCGGTCACT |
|
R: CATACGACTGGTCAGAGTCAGG |
||
|
26 |
RNF168 |
F: GCCAGTTCGTCTGCTCAGTA |
|
R: CTGCCGCCACCTTGCTTAT |
||
|
27 |
HIST1 |
F: AAGAGCCTGGTAAGCAAGGG |
|
R: TGCACCCGTTGCCTTAGTTT |
||
|
28 |
TopBP1 |
F: CCAACGAGTTCAGAAATGTCCAG |
|
R: AACGCCACTAAAAGGGTCACA |
||
|
29 |
ACTB |
F: AACCGCGAGAAGATGACCC |
|
R: AGCACAGCCTGG TAGCAAC |
||
|
30 |
H2AX |
F: GGCCTCCCAGGAGTACTAAGA |
|
R: CTCTTTCCATGAGGGCGGTG |
||
|
31 |
XRCC4 |
F: TTGATCTGTGAAAGCGGGCG |
|
R: TCTCCATTTCTTAATACCTCTCCGT |
||
|
32 |
RBBP8 |
F: CGAGGATTTGGCACTCTGGT |
|
R: ACAGGTCAAATACCGCCTCC |
||
|
33 |
EP300 |
F: GCCCTCTACCTGACCCAAGT |
|
R: ATAGCCCATAGGCGGGTTG |
||
|
34 |
LIG4 |
F: GGCTTGACGTCAGGAAACCAT |
|
R: GAAGTTTGTGAGGCAGCCAT |
||
|
35 |
C-FLIP |
F: GTGACAGCTGAGACAACAAGG |
|
R: TGGGGGAGTTCGTCCTGT |
Примечание: F – прямой праймер, R – обратный праймер
Постановку каждого локуса проводили в трех технических повторах. Для амплификации использовали реакционную смесь, содержащую 1х PCR буфер, 1,5 мМ MgCl2, смесь 0,2 мМ dNTP, по 500 нМ праймеров, 0,05 U/мкл Taq-полимеразы, 10 нг кДНК. Для внутреннего контроля использовали NTC и NRT пробы. Полученные смеси амплифицировали в термоциклере CFX 96 (Bio-Rad, США) в соответствии с температурным режимом: t=95°С – 240 сек; 40 циклов: t=95°С – 10 сек, t=58°С – 30 сек, t=72°С – 30 сек.
Относительную экспрессию генов (RЕ) рассчитывали по формуле RЕ=2-ΔΔCt. Нормализацию результатов проводили по 3 референсным генам (GAPDH, ACTB и B2M) и уровню экспрессии соответствующих генов-мишеней в образцах контрольной группы, последовательно по схеме, приведенной ниже.
1. Нормализация по среднему геометрическому референсных локусов: ΔC(t)=C(t)target–среднее геометрическоеC(t)reference.
2. Расчет среднего арифметического ΔC(t) по каждому гену для контрольной (интактной) и облученной групп.
3. Нормализация по контрольной (интактной) группе: ΔΔC(t)= среднее арифметическоеΔC(t)облученной группы – среднее арифметическоеΔC(t) контрольной группы.
4. Окончательный результат (кратное различие): 2-ΔΔC(t).[6].
Статистическую обработку данных проводили с применением однофакторного дисперсионного анализа (One-Way ANOVA) в программе Statistica 8.0 (StatSoft Inc.). Для проведения кластерного анализа (Hierarchical Clustering, Euclidean distance) и построения тепловых карт (heat map) использовали собственные скрипты на языке R (R-Studio 8.10.173.987). Для построения диаграмм Эдвардса–Венна применяли алгоритм сравнения большего числа множеств, реализованный на JavaScript. Кластеризацию генов по функции проводили с помощью алгоритма FMD (Functional module detection). Значение Q каждого члена, связанного с функциональными модулями, рассчитывали с использованием одностороннего точного критерия Фишера и поправки Бенджамини–Хохберга для корректировки множественного сравнения [7].
Результаты исследования и их обсуждение
Ионизирующее излучение вызывает двухцепочечные разрывы в ДНК опухолевых клеток, но некоторые клоны опухолевых клеток обладают способностью повышать активность системы репарации двухцепочечных разрывов для преодоления цитотоксичности от излучения, а также иных сигнальных путей, не являющихся компонентами репарационных систем клетки [8].
После нашего модельного эксперимента только определенный пул клеток НТ-29 сохранил жизнеспособность. Для дозы 5 Гр доля таких клеток составила 32%, а для дозы 7 Гр – порядка 20% от изначального количества клеток, взятых в эксперимент. Данный эффект может быть связан с повышенной устойчивостью этих клеток к облучению, сформировавшейся благодаря определенным молекулярно-генетическим особенностям. Так, в результате эксперимента по облучению клеток HT-29 были получены данные о дифференциальном показателе экспрессии 32 генов в интактных (контрольных) и облученных клетках (рис. 2).
По особенностям экспрессии в интактных и облученных образцах было выделено 3 основных кластера генов: 1 – BRCA2, C-FLIP, CASP3, XRCC4, KU70, CDK1, 2 – AKT, RBBP8, RAP80, EP300, BCL2, RAD50, FGFR2, H2AX, CCND3, CCND1 и 3 – TOPB1, ATM, RNF168 PTEN, BRIP, EXO1, BAX, TP53, CASP9, RIF1, CASP8, BRCA1, CDKN1B, HIST1, LIG4.
При этом сами интактные и облученные группы клеток по идентичности экспрессии генов объединяются в кластеры (рис. 2, 3). Сходство по уровню экспрессии некоторых генов в интактных и облученных клетках на разных уровнях кластеризации (рис. 2, 3 и табл. 2) можно объяснить изначальным наличием в линии НТ-29 клонов с определенным уровнем экспрессии рассматриваемых генов, который в условиях облучения обеспечил их селективное выживание.
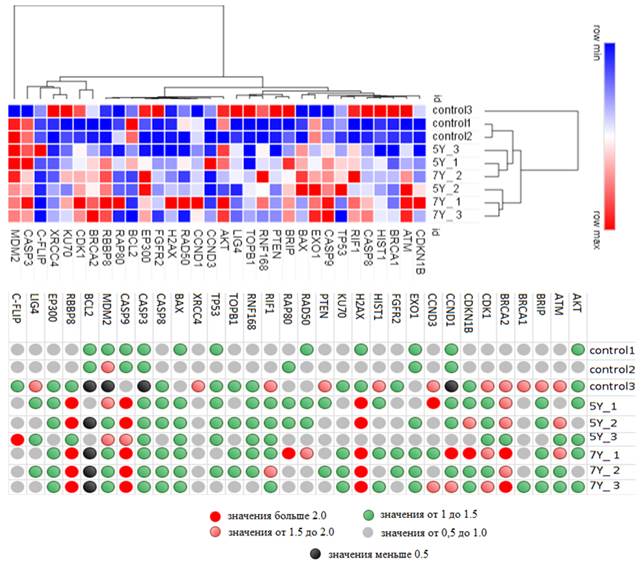
Рис. 2. Heatmap и кластерный анализ дифференциальной экспрессии 32 генов в интактных (контрольных) и облученных клетках НТ-29
Таблица 2
Сходства и различия в показателе экспрессии генов между кластерами
|
Ген |
Количество групп, в которых повышена экспрессия |
Наименование групп |
|
BRCA2 |
6 |
control3, 5Y_1, 5Y_2, 7Y_1, 7Y_2, 7Y_3 |
|
CASP9 |
6 |
5Y_1, 5Y_2, 5Y_3, 7Y_1, 7Y_2, 7Y_3 |
|
H2AX |
5 |
5Y_1, 5Y_2, 7Y_1, 7Y_2, 7Y_3 |
|
RBBP8 |
5 |
5Y_1, 5Y_2, 7Y_1, 7Y_2, 7Y_3 |
|
ATM |
3 |
control3, 5Y_2, 7Y_1 |
|
CCND3 |
3 |
control3, 5Y_1, 7Y_3 |
|
CDK1 |
3 |
control3, 7Y_1, 7Y_3 |
|
MDM2 |
3 |
control2, 5Y_1, 5Y_3 |
|
CCND1 |
2 |
7Y_1, 7Y_3 |
|
CDKN1B |
2 |
5Y_2, 7Y_2 |
|
RIF1 |
2 |
control3, 7Y_2 |
Стоит отметить, что 4 гена (BRCA2, CASP9, H2AX, RBBP8) имели повышенную экспрессию в более чем в 5 исследуемых группах, при этом повышенная экспрессия гена BRCA2 наблюдалась и у части интактных клеток (табл. 2).
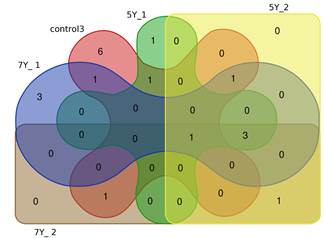
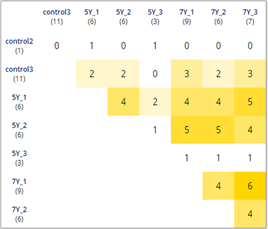
Рис. 3. Диаграмма Эдвардса–Венна (показано сходство по количеству генов с повышенной экспрессией в 9 группах – 3 контрольных, 3 облученных в дозе 5 Гр и 3 – в дозе 7 Гр)
После нормализации показателей относительной экспрессии в клетках, подвергнутых облучению 5 и 7 Гр, были получены следующие данные: статистически значимо была повышена экспрессия гена RBBP8 – в 1,7 и 2,2 раза, BRCA2 – в 1,7 и 2,3 раза, H2AX – в 2,3 и 4,5 раза и CASP9 – в 2,0 и 2,4 раза соответственно, снижена экспрессия гена BCL2 – в 1,6 и 3,0 раза (р<0,05) соответственно относительно интактных клеток. В клетках, подвергнутых облучению 7 Гр, также наблюдалось статистически значимое повышение экспрессии гена RIF1 – в 1,5 раза (р<0,05) (рис. 4).
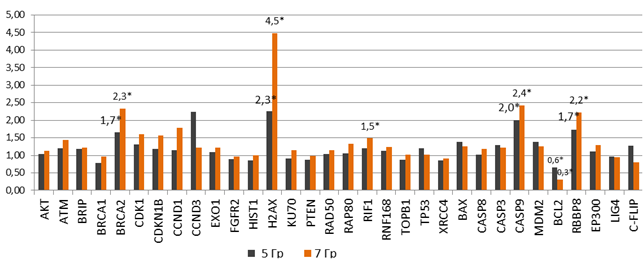
Рис. 4. Относительная экспрессия генов в клетках НТ-29 с повышенной радиорезистентностью (после облучения в дозах 5 и 7 Гр в течение 5 дней)
*– статистически значимые отличия относительно интактных (контрольных) клеток (р<0,05)
С помощью алгоритма FMD исследованные нами гены ATM, AKT, BRCA1, BRCA2, BRIP, CDK1, CDKN1B, CCND1, CCND3, FGFR2, HIST1, H2AX, KU70, EXO1, PTEN, RAD50, RAP80, RNF168, TOPB1, RIF1TP53, MDM2, XRCC4, BAX, BCL2, CASP8, CASP3, CASP9, RBBP8, EP300, LIG4, C-FLIP были разделены на 6 функциональных модулей и визуально представлены на рисунке 5.
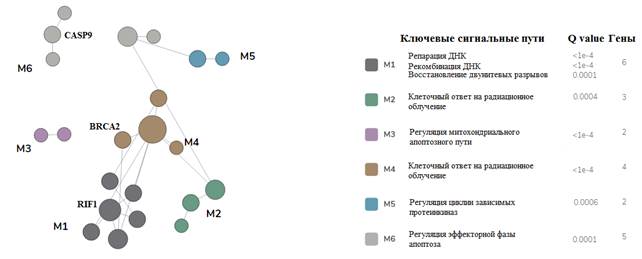
Рис. 5. Функциональная классификация сигнальных путей, включающих 32 генетических локуса
Из данных, представленных на рисунке 5, видно, что изменение экспрессии генов BRCA2, CASP9 и RIF1 затрагивает 3 функциональных модуля, объединяющих группу сигнальных клеточных каскадов, таких как регуляция апоптоза (М6, Q=0,0001), клеточный ответ на радиационное облучение (М4, Q< 1e-4) и репарация ДНК (М1, Q< 1e-4).
Применение алгоритма «сетевой интеграции с несколькими ассоциациями» позволило установить взаимодействия между генами BCL2, BRCA2, H2AX, CASP9, RBBP8, RIF1 и APAF1 (apoptotic peptidase activating factor 1), SPO11 (initiator of meiotic double stranded breaks), PALB2 (partner and localizer of BRCA2), APPL1 (adaptor protein, phosphotyrosine interacting with PH domain and leucine zipper 1), PSIP1 (PC4 and SFRS1 interacting protein 1), DIABLO (diablo IAP-binding mitochondrial protein), DMC1 (DNA meiotic recombinase 1) BIK, (BCL2 interacting killer), BAZ1B (bromodomain adjacent to zinc finger domain 1B), FKBP8 (FK506 binding protein 8), BAG1 (BCL2 associated athanogene 1), RAD51, VDAC1 (voltage dependent anion channel 1), XRCC3 (X-ray repair cross complementing 3), DCC (DCC netrin 1 receptor) RAD50 (RAD50 double strand break repair protein), PSMC3IP (PSMC3 interacting protein), TEX15 (testis expressed 15), MND1 (meiotic nuclear divisions 1) и AIFM1 (apoptosis inducing factor, mitochondria associated 1) (рис. 6). Данный алгоритм предсказывает функцию гена в составе сложной сети из множества генов посредством использования одого из методов машинного обучения, основанного на распределении меток гауссовых полей. Алгоритм присваивает оценку каждому узлу-точке построенной сети, отражающую вычисленную силу связи [9]. Соответственно BRCA2, H2AX, BCL2, CASP9 и RBBP8 являются компонентами различных сигнальных путей опухолевых клеток, изменение их транскрипционой активности приводит к модуляции активности целого ряда других генов (рис. 6).
Так, в ответ на двухцепочечные разрывы в ДНК, вызванные ионизирующим излучением, H2AX (гистоновый белок, кодируемый геном H2AX) становится фосфорилированным по серину (γH2AX). Из-за этого ДНК становится менее конденсированной, и освобождается место для присоединения белковых комплексов, например MRN (состоящего из Mre11, Rad50 и Nbs1), RAD51 и АТМ), необходимых для реализации процесса репарации [10, 11].
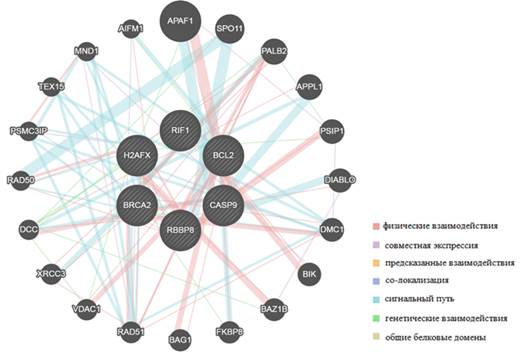
Рис. 6. Взаимодействия генов, вычисленные с помощью алгоритма сетевой интеграции с несколькими ассоциациями
Белок, кодируемый геном BRCA2, также необходим для восстановления поврежденной излучением ДНК (напрямую взаимодействует с рекомбиназой RAD51, стимулируя важный этап гомологичной рекомбинации) [12]. RBBP8 кодирует белок, который регулирует пролиферацию клеток [13]. Соответственно повышенная экспрессия генов BRCA2, H2AX и RBBP8 в отдельных клонах клеточной линии НТ-29, вероятно, обеспечивает большую эффективность системы репарации ДНК, что дает им преимущество и обеспечивает выживание при воздействии лучевой терапии.
Важная роль в регуляцию апоптоза отведена белкам BCL2 и CASP9. BCL2 подавляет апоптоз, изменяя проницаемость митохондриальной мембраны, что предотвращает выход цитохрома C из митохондрий и ингибирует каспазы (также ингибирование реализуется за счет связывания BCL2 – фактора, активирующего апоптоз APAF1). Наоборот, инициаторная каспаза 9 является критическим белком для запуска апоптоза, который активируется APAF1 путем расщепления про-каспазы-9 [14]. Соответственно повышенная экспрессия CASP9 и сниженная экспрессия BCL2 могут способствовать более эффективному протеканию процесса апоптоза, чем в интактных клетках. Можно предположить, что данная особенность способна оказывать определенное регулирующее воздействие на численность клонов НТ-29, обладающих описанными выше молекулярно-генетическими особенностями.
Заключение
Таким образом, данное исследование позволило установить исходную гетерогенность клеток НТ-29 по транскрипционной активности ряда генов, регулирующих систему репарации и апоптоз. Эта гетерогенность лежит в основе селективного выживания клеток в условиях лучевой терапии при 5 и 7 Гр: выжившие клетки обладают более эффективной системой репарации ДНК (повышенная транскрипционная активность генов BRCA2, H2AX, RBBP8) и более эффективной системой регуляции апоптоза (повышенная транскрипционная активность проапоптозного гена CASP9 и сниженная транскрипционная активность антиапоптозного гена BCL2).
Исследование выполнено в рамках гос. задания «Поиск предикторов радиорезистентности рака прямой кишки и разработка персонифицированных неоадъювантных терапевтических подходов».
Библиографическая ссылка
Кутилин Д.С., Гусарева М.А., Кошелева Н.Г., Габричидзе П.Н., Донцов В.А., Легостаев В.М., Шляхова О.В., Лиман Н.А., Солнцева А.А., Крохмаль Ю.Н. АБЕРРАНТНАЯ ТРАНСКРИПЦИОННАЯ АКТИВНОСТЬ ГЕНОВ КАК ФАКТОР РАДИОРЕЗИСТЕНТНОСТИ КЛЕТОК ЛИНИИ HT-29 // Современные проблемы науки и образования. 2020. № 3. ;URL: https://science-education.ru/ru/article/view?id=29831 (дата обращения: 01.07.2025).
DOI: https://doi.org/10.17513/spno.29831