



Эндопаразитические эвгленовые жгутиконосцы, относящиеся к отряду Parastasiida Likhaschev, 1998 (Euglenoidea, Euglenozoa) к родам Parastasia Michajlow, 1972 и Sophiensia Likhaschev, 1998 в качестве хозяев которых известны пресноводные беспозвоночные различных систематических групп, по настоящее время изучены недостаточно [1, 6, 7]. Для России фауна парастазиид изучалась в водоемах Ленинградской, Омской, Мурманской и Московской областей, в водоемах других регионов видовой состав этих жгутиконосцев остается неизвестным [1, 9]. Мало изученной остается биология парастазиид, сезонная изменчивость в жизненных циклах. Недостаточно изучена морфология отдельных видов парастазиид и ее изменения под влиянием сезонного и географического факторов. Нет подробных данных о репродуктивной фазе жизненных циклов для целого ряда видов. В литературе практически нет данных о влиянии факторов среды на жизненные циклы парастазиид, отсутствуют сведения об их взаимоотношениях с хозяевами в системе «паразит – хозяин», нет оценки их значения в системе пресноводного биоценоза. Большинство известных видов этих жгутиконосцев живет в пищеварительном тракте, яйцевых мешках и полости тела пресноводных циклопид (Cyclopidae, Copepoda) [1, 2, 10]. Несколько видов встречено в организмах инфузорий, турбеллярий, нематод, коловраток, гастротрих и олигохет. Эти виды эвгленовых жгутиконосцев отнесены авторами к роду Astasia, описаны давно и в последние годы о них не появлялось новых данных. Поскольку парастазииды – агамно размножающиеся простейшие, важно изучение морфофизиологических адаптаций их видов и внутривидовой изменчивости для установления возможных путей эволюции этой группы эвгленид [6, 7].
Материал и методы исследования
Материал собирался в разнотипных водоемах г. Омска и г. Ишима (Тюменская область) в период с 2003 по 2015 г. За данный период было собрано около 1 тыс. проб. Сбор и обработка полевого материала проводилась с использованием стандартных гидробиологических, цитохимических и протозоологических методик [4, 5, 6, 8]. Определение видовой принадлежности найденных форм проводилось на живом материале при увеличении ок. х15, об. х40 и с использованием фазово-контрастного устройства. Изучение жизненных циклов парастазиид проводили в микроаквариумах по описанной в литературе методике [1, 8, 10]. Видовое определение приводилось по описаниям, содержащимся в литературе [2, 3, 4, 10].
Результаты исследования и их обсуждение
Жизненный цикл парастазиид включает 2 стадии – трофическую и репродуктивную. Трофическая стадия, проходящая в организме хозяина, заключается в росте трофозоита (питающейся особи) и накоплении им парамилона. Репродуктивная стадия, проходящая в воде, заключается в последовательных палинтомических делениях и образовании многочисленных инвазионных жгутиковых клеток (зооспор) (рис. 1).
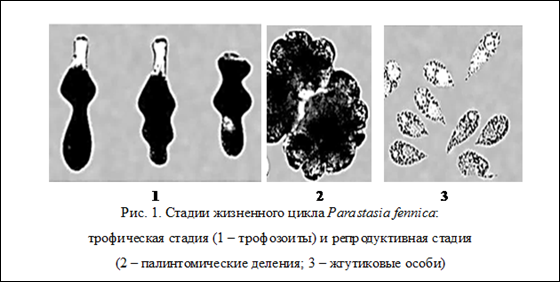
Трофозоиты всех известных видов парастазиид обладают специфической формой движения, которая выражается в метаболических сокращениях тела соответствующих по направлению перистальтике кишечника хозяина (рис. 1.1). Метаболические сокращения клетки имеют адаптивный характер, предотвращая выведение незрелых (не готовых к делению) трофозоитов во внешнюю среду и, способствуя выходу из кишечника зрелых особей, готовых к палинтомическим делениям. Характер движения трофозоитов зависит от строения стенки клетки, образованной пелликулой.
Пелликула трофозоитов плотная, снабженная большим числом наружных ребер, образующих косую штриховку на поверхности клетки. По своему строению пелликула трофозоита, например вида Parastasia fennica, типична для эвгленовых жгутиконосцев. Она образована плазмалеммой, состоящей из цитоплазматической мембраны с хорошо развитым слоем гликокаликса. В кортикальном слое цитоплазмы с плазматической мембраной ассоциированы комплексы микротрубочек и микрофиламентов, имеющие, по-видимому, опорное значение. У основания пелликулярных ребер находятся экструсомы – мукоцисты (слизеносные тельца).
В цитоплазмена ходятся митохондрии с пластичатыми кристами. Аппарат Гольджи представлен несколькими диктиосомами. Гранулярнный и агранулярный эндоплазматический ретикулум также хорошо развит. Жгутиковый аппарат парастазиид состоит из двух жгутиков неравной длины, но оба жгутика не выходят за пределы жгутикового резервуара. Короткий жгутик снабжен паракристаллиновыми структурами, характерными для примитивных видов эвгленовых жгутиконосцев.
Важным родовым признаком парастазиид служит наличие/отсутствие стигмы. Виды рода Parastasia стигмы не имеют, к ним относятся: Parastasia fennica, P.coelomae, P.cyclops и др. Стигму имеют все виды рода Sophiensia: S. macrogranulata, S. caudata, S. ucrainica, S. hanoiensis.
В цитоплазме много первичных и вторичных лизосом. Пиноцитоз, вероятно, осуществляется всей поверхностью клетки, включая и плазматическую мембрану жгутикового резервуара.
Основным резервным веществом трофозоита является парамилон, накапливающийся в виде многочисленных гранул/зерен различного размера и формы у разных парастазиид. У большинства видов парастазиид парамилоновые зерна довольно мелкие, достигают в длину 1–2,5 мкм и имеют округлую или палочковидную форму. Только у Sophiensia macrogranulata зерна парамилона крупные, достигают в длину 3–5 мкм.
Для трофозоитов характерны большие запасы зерен парамилона, которые заполняют практически всю эндоплазму и перемещаются из одной части клетки в другую при метаболических сокращениях клетки.
Кроме парамилона у парастазиид в качестве резервного вещества откладывается гликоген. Запасы его невелики, зерна мелкие и располагаются между зернами парамилона. Наличие этого полисахарида у парастазиид указывает на то, что они перешли на гетеротрофный обмен веществ и по характеру клеточного метаболизма отличаются от фотосинтезирующих видов эвгленовых жгутиконосцев.
Трофозоиты всех известных в настоящее время видов парастазиид имеют одно округлое ядро с центральным ядрышком. Диаметр ядра различен у разных видов и зависит от размеров клетки. В организме хозяина трофозоиты всех известных видов парастазиид не делятся, их ядро находится в интерфазном состоянии и расположено ближе к заднему концу клетки.
Анализ наших и литературных данных показывает, что трофозоиты всех известных видов рода Parastasia обнаруживают общность организации клетки [1, 10]. Их трофозоиты имеют также ряд общих особенностей морфологии и физиологии с видами рода Astasia: пелликула, жгутиковый резервуар, жгутики, ядро, парамилон, форма движения, способ питания. В свою очередь виды рода Sophiensia обнаруживают общность организации как с родом Astasia, так и с родом Khawkinea, лишь виды которого среди всех свободноживущих бесцветных эвгленовых жгутиконосцев имеют стигму.
Репродуктивная фаза жизненного цикла парастазиид начинается с выхода трофозоита из организма хозяина в воду. Следует отметить, что готовность трофозоита к репродуктивной стадии, а следовательно, и к выходу в воду определяются размерами трофозоита, объемом запасов парамилона и миграцией ядра к передней части клетки.
Размножение парастазиид подробно изучено в лабораторной культуре. В микроаквариуме трофозоит, вышедший из организма хозяина, движется по субстрату с помощью метаболических сокращений, а затем у одних видов парастазиид (S. caudata, S. macrogranulata и др.) вышедший трофозоит образует плавательный жгутик, и они начинает плавать в воде. У других видов (P. fennica и др.) жгутик после выхода трофозоита в воду не образуется, и особь ползает по субстрату.
Время существования трофозоита в воде у разных видов парастазиид неодинаково и в большей степени зависит от внешних условий. После периода свободного плавания в воде или движения путем метаболических сокращений тела трофозоит прикрепляется к субстрату и начинает делиться (рис. 1.2.). Основная форма агамного размножения парастазиид – палинтомия, которая свойственна большинству видов парастазиид и заключается в последовательных делениях сначала материнской особи, а затем и дочерних клеток, без роста этих клеток и с образованием в конечном итоге большого числа инвазионных жгутиковых клеток (зооспор). Однако В. Михайлов [10] считает, что кроме палинтомии у парастазиид есть и другая форма деления – синтомия. Под синтомией понимается процесс образования большого числа ядер в материнской клетке, вокруг которых обособляется цитоплазма и формируются дочерние клетки под материнской оболочкой. Синтомия заканчивается образованием большого числа инвазионных особей очень мелких размеров (зооспоры).
Репродуктивная фаза заканчивается формированием инвазионных жгутиковых (зооспоры) (рис. 1.3.). Эти клетки образуются в результате последнего палинтомического или синтомического деления, и число их различно у разных видов парастазиид.
Наибольшее число зооспор характерно для трех видов: P. coelomae, P. bulgarica, в результате синтомии, у трофозоитов которых образуется 200–500 клеток. У P. helwetica и P. granulosa при палинтомии образуется не более 4 жгутиковых клеток. В среднем у большинства видов парастазиид образуется от 16 до 128 жгутиковых клеток.
Жгутиковые зооспоры имеют целый ряд особенностей морфологии, которые отличают их от трофозоитов, хотя основные морфологические черты, свойственные трофозоитам, у них сохраняются. Жгутиковые зооспоры имеют размеры в 5 и более раз меньше размеров трофозоитов.
У всех видов парастазиид зооспоры имеют жгутик, с помощью которого они движутся в воде, а если такая клетка прикрепляется к субстрату, то жгутик продолжает находиться в движении, способствуя, вероятно, привлечению рачков, использующих зооспоры в пищу и заражающимися парастазиидами. У многих видов парастазиид жгутиковые зооспоры имеют прикрепительную ножку или «стебелек», с помощью которого и осуществляется прикрепление клеток к субстрату.
Жгутиковые зооспоры характеризуются незначительными запасами парамилона, и если они через несколько дней не попадают в кишечник хозяина, то погибают. Вероятно, жгутиковые зооспоры не питаются и расходуют для жизнедеятельности запасы парамилона.
Краткое рассмотрение морфофизиологических особенностей всех стадий жизненного циклапарастазиид позволило выявить адаптации, которые являются наиболее существенными маркерами эволюции паразитизма у этой группы эвгленовых жгутиконосцев:
1.Трофозоит, запасающий парамилон;
2. Палинтомия, в результате которой формируется большое число дочерних особей;
3. Жгутиковые клетки (зооспоры), имеющие инвазионные морфологию и поведение.
Эти адаптации сформировались в результате длительной эволюции через трофическую систему «хищник – жертва» = «циклоп – гипотетический свободноживущий предок парастазиид» к системе «паразит – хозяин» = «циклоп – парастазииды».
Эвглениды, подобные видам рода Euglena, дали начало бесцветным астазиидам. Некоторые из них в процессе морфологической эволюции почти не изменились с тех пор, как приобрели хлоропласты и вторично потеряли их, сохранив стигму и парамилон, что характерно для видов рода Khawkinea. Другие, в частности, виды рода Astasia, при очень высокой степени сходства с окрашенными эвгленовыми утратили не только хлоропласты, но и стигму. Некоторые астазииды, перейдя к паразитизму в организмах водных беспозвоночных, дали начало видам родов Parastasia и Sophiensia. В. Михайлов считает, что парастазии происходят от свободноживущих эвгленовых, подобных астазиевым [10]. В связи с тем, что парастазииды эволюционно молодые паразиты, то они не имеют значительных отличий от свободноживущих бесцветных эвгленовых. Эволюция их шла по линии приспособления к обитанию в организме рачка-хозяина, а также к делению в воде. В процессе эволюции, в жизненном цикле парастазиид возникли адаптации, которые тесно связывают их с жизненными циклами хозяев [1, 10]. Парастазииды рано отделились в качестве самостоятельной группы от анцестральных форм, на что указывает обнаружение у них дополнительного жгутика с паракристаллиновыми структурами, сходными с таковыми у примитивных форм Euglenidae [7]. Клеточный метаболизм парастазиид, хотя и типичен для эвгленовых жгутиконосцев в целом, но имеет некоторые особенности. К числу их относится преобладание ферментов гликолитического цикла (лактатдегидрогеназа и др.) над активностью основных ферментов цикла Кребса (сукцинатдегидрогеназа и др.) на стадии трофозоита, а на репродуктивной стадии наоборот [1].
Возможно, предположить алиментарный путь возникновения и эволюции облигатного паразитизма у парастазиид. Их анцестральные формы были свободноживущими бесцветными эвгленовыми (у имеющих стигму софиензий подобные видам рода Khawkinea, у парастазий, подобные видам рода Astasia). Предковые свободноживущие формы входили в пищевой спектр циклопид. Они подобно многим другим простейшим, в качестве пищевого объекта, попав в кишечник рачков, либо переваривались, либо проходили транзитом. Некоторые формы парастазиид, проходя транзитом через пищеварительный тракт рачков, задерживались и, будучи сапрофитами, питались продуктами пищеварения хозяина. В результате эти формы перешли к факультативному паразитизму. После приобретения комплекса морфофизиологических и биохимических адаптаций к жизни в организме рачка парастазииды становятся облигатными паразитами.
Возможно, в процессе длительной эволюции вначале возникли и существовали какие-то промежуточные формы, которые были связующим звеном между исходными окрашенными и современными вторично бесцветными видами, о которых мы сейчас ничего не знаем. Поэтому, вероятно, о прямом филогенетическом родстве между родами эвгленовых жгутиконосцев можно говорить лишь в некоторых случаях, например в линиях: Euglena – Khawkinea – Astasia (окрашенные и вторично бесцветные роды) и Khawkinea – Astasia – Sophiensia – Parastasia (свободноживущие и паразитические вторично бесцветные роды).
Библиографическая ссылка
Лихачев С.Ф., Тавадзе Б.Д., Монтина И.М. МОРФОЛОГИЧЕСКИЕ И ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ В ЭВОЛЮЦИИ ПАРАСТАЗИИД (PARASTASIIDA, EUGLENOZOA) // Современные проблемы науки и образования. 2016. № 2. ;URL: https://science-education.ru/ru/article/view?id=24314 (дата обращения: 04.07.2025).