



Процесс развития периферической нервной системы в филогенезе отмечается большой динамичностью, в связи с этим выявление определенных закономерностей и параллелей имеет не только теоретический интерес, но представляет собой большую практическую ценность. Еще в 1966 году Н. И. Зазыбин, в результате проведенного микроморфологического исследования, убедительно доказал, что «…нервная система представляет собой активный материал и существенно влияет на развитие других тканей и органов» [9.10].
Таким образом, целью нашего исследования явилось изучение литературных источников по проблеме образования и особенностей развития периферических нервов плечевого сплетения в эволюционном ряду.
Вначале – небольшой экскурс в историю. Изучением анатомических особенностей периферических нервов передней конечности животных и верхней конечности человека занимались ученые еще в глубокой древности. Благодаря знаменитому римскому врачу Галену, в 131–211 г. н. э. сравнительное изучение животных достигло значительного развития. В XVI веке в работах Андрея Везалия и Леонардо да Винчи анатомические знания о плечевом сплетении получили дальнейшее право на существование. В конце 19 века были изучены длинные ветви плечевого сплетения у человека и ряда животных. Особенно интересны работы В. Грубера (1870), который, изучая строение нервов верхней конечности, подробно описал связи между срединным и локтевым нервами в области предплечья. Интересно отметить, что все исследователи этого периода устанавливали частные варианты в строении, но не могли их систематизировать и представить формы изменчивости в анатомии нервов верхней конечности. Только Н. И. Пирогов (1843–1845) в ряде своих работ указывал на необходимость изучения индивидуальной изменчивости в строении систем и органов. В 90-х годах 19 века и в начале 20 века благодаря новым методам исследования начали изучать внутриствольную топографию нервов верхней конечности. Изучением анатомического строения плечевого сплетения и его длинных ветвей занимались многие авторы [2,12]. Литературные данные показывают, что нервы верхней конечности имеют очень изменчивое строение [7,12]. Такую изменчивость в строении большинство современных авторов объясняет общностью происхождения в филоонтогенезе (Борисов И. Н., 1986; Асатиани Д. Л., 1989). По современным представлениям И. В. Гайворонского (2011), С. С. Михайлова (2011), И. П. Горшкова (2009), формирование плечевого сплетения начинается из спинномозговых нервов (С5 - Th1 и частично Th2) в результате их дихотомии на передние и задние ветви [8,11]. Передние ветви группируются в три первичных пучках (верхний, средний и нижний), которые в свою очередь разделяются (вторая дихотомия) на передние и задние ветви. Затем идет образование трех вторичных пучков: наружный, внутренний и задний, расположенных в подключичном пространстве вокруг подмышечной артерии. Далее, дорсальные ветви первичных пучков образуют задний вторичный пучок. Вентральные ветви от первичных пучков формируют два вторичных пучка: наружный и внутренний. При этом вторичный наружный пучок образуется путем объединения передних ветвей верхнего и среднего первичных пучков плечевого сплетения. Вторичный внутренний пучок образуется передней ветвью первичного нижнего пучка, которая в образовании наружного и заднего вторичного пучка участия не принимает. На этом этапе «деления – объединения» формирование плечевого сплетения заканчивается (рисунок 1А). Авторы (М. Р. Сапин, Д. Б. Никитюк, 2001; М. Г. Привес, Н. К. Лысенков, В. И. Бушкович, 1985), изучавшие сегментарный состав периферических нервов, указывают на большую вариабельность происхождения этих стволов. Например, срединный нерв формируется чаще из C6 - Th1 или C5 - Th1, локтевой – из C7, C8, Th1 или C8, Th1, лучевой – из C5 - C8- Th1 или C6 - C7, мышечно-кожный – из С5 - С6 или С5 - С7, Th1.
А.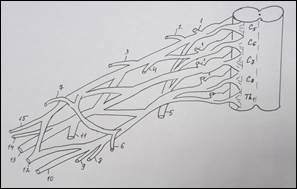 Б.
Б.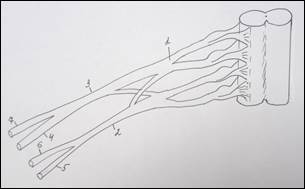
В.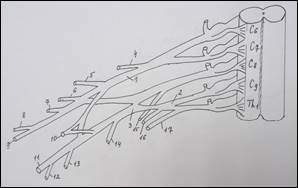 Г.
Г.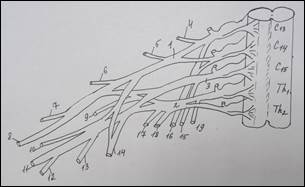
Д.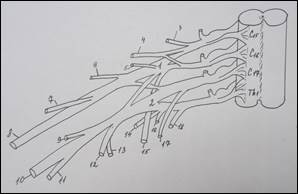 Е.
Е.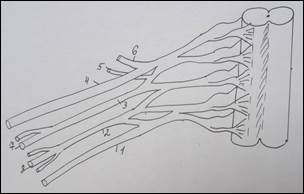
Схема строения плечевого сплетения у человека (А):
1 – n. culaneus brachii med.; 2 — n. cutaneus antebrachii medialis; 3 – n. ulnaris; 4 – n. radialis; 5 – n. medianus; 6 – n. axillaris; 7 – n. musculocutaneus; 8 – fasciculus lat.; 9 – n. suprascapularis; 10 – fasciculus post.; 11 – n. thoracicus longus; 12 – fasciculus med; 13 – n. cruris; 14 – n. iliohypogastricus; 15 – n. spinalis. У земноводных (Б): 1 – краниальный ствол, 2 – каудальный ствол, 3 – каудальный вторичный; 4 – нерв разгибателей, 5 – мышечно-кожный, 6 – нерв сгибателей, 7 – подмышечный. У пресмыкающихся (В): 1 – краниальный ствол, 2 – каудальный ствол, 3 – каудальный вторичный, 4 – надлопаточный, 5 – краниальный грудной, 6 – лопаточно – плечевой, 7 – надкоракоидный, 8 – подмышечный, 9 – нерв разгибателей, 10 – медиальный кожный, 11 – нерв сгибателей, 12 – мышечно-кожный, 13 – каудальный грудной, 14 – коракоидно-плечевой, 15 – длинный грудной, 16 – надкоракоидный, 17 – предлопаточный. У голубя (Г): 1 – краниальный ствол, 2 – каудальный ствол, 3 – средний ствол, 4 - дорсальный лопаточный, 5 – надкоракоидный, 6 – краниальный грудной, 7 – подмышечный, 8 – нерв разгибателей, 9 – подлопаточный, 10 - медиальный кожный, 11 – нерв сгибателей, 12 – мышечно-кожный, 13 – коракоидно-плечевой задний, 14 – коракоидно – плечевой передний, 15 – дорсальный грудной, 16 – длинный грудной, 17 – каудальный грудной, 18 – лопаточно-плечевой, 19 – предлопаточный. У курицы (Д): 1 – краниальный ствол, 2 – каудальный ствол, 3 – дорсальный лопаточный, 4 – надкоракоидный, 5 – краниальный грудной, 6 – подлопаточный, 7 – подмышечный, 8 – нерв разгибателей, 9 –медиальный кожный, 10 – нерв сгибателей, 11 – мышечно-кожный, 12 – коракоидно-плечевой задний, 13 – коракоидно-плечевой передний, 14 – дорсальный грудной, 15 – длинный грудной, 16 – лопаточно-плечевой, 17 – каудальный грудной, 18 – предлопаточный. У млекопитающих (Е): 1 – локтевой нерв, 2 – срединный нерв, 3 – лучевой нерв, 4 – мышечно-кожный нерв, 5 – подмышечный нерв, 6 – передний грудной нерв, 7 – поверхностная ветвь лучевого нерва, 8 – глубокая ветвь лучевого нерва.
В исследованиях ряда авторов указаны крайние формы изменчивости нервных стволов верхней конечности. Крайние формы изменчивости мышечно-кожного нерва были представлены еще в работах К. А. Григоровича (1946), В. П. Аратского (1950), А. Н. Микитюка (1983), которые различали рассыпанную (сетевидную) и магистральную (концентрированную) формы этого нерва. К. А. Григорович выделял форму изолированного ствола мышечно-кожного нерва и форму, когда этот нерв замещен ветвями срединного. Следует отметить, что классификация крайних форм изменчивости нерва более согласуется с филогенезом этого нерва. Ряд авторов придает большое значение уровню формирования ствола срединного нерва. Так, В. Н. Шевкуненко (1949), Л. В. Некрасова (2011), Ю. П. Турсунова (2011) на основании данных сравнительной анатомии показал, что срединный нерв выше всего формируется из своих корешков у человека [15]. В образовании локтевого нерва участвует меньшее число сегментных нервов, чем в образовании срединного. Поэтому он варьирует в своей структуре в меньшей степени, чем срединный (Bowen B. C., 2004). Что касается сравнительно-анатомических исследований морфологических особенностей плечевого сплетения и уточнения уровня его формирования у различных представителей позвоночных животных, следует остановиться лишь на основных выводах из работ главным образом современных исследователей. Наиболее распространенным критерием оценки степени развития плечевого сплетения принято считать количество спинномозговых нервов, участвующих в его формировании. Анализируя данные литературы, напрашивается вывод о том, что грудные конечности позвоночных животных иннервируются производными плечевого сплетения, которое обычно образуется вентральными ветвями нескольких спинномозговых нервов. Исключение составляют некоторые бесхвостые земноводные (жабы), у которых плечевое сплетение, по существу, отсутствует, и иннервация ее грудных конечностей осуществляется ветвями одного спинномозгового нерва [14]. Схожая картина наблюдается и у лягушек, хотя по данным А. Д. Ноздрачева (1994), Е. Л. Полякова (1994), Н. В. Балеева (2009) в формировании плечевого сплетения у земноводных участвуют 2–3 нерва, а именно – II, III и IV краниальные спинномозговые нервы (рисунок 1Б). У рептилий, в связи с выходом на сушу и большей сложностью движений, плечевое сплетение организуется сложнее. Для него уже характерно разделение сплетения на шейное и плечевое. По данным М. В. Веселовского и Г. И. Василевской (2001) анатомическое строение плечевого сплетения асимметрично. На левой конечности плечевое сплетение образовано четырьмя спинномозговыми нервами: двумя последними шейными и двумя первыми грудными. Шейные спинномозговые нервы объединяются в короткий общий ствол. Грудные нервы проходят самостоятельно до уровня капсулы плечевого сустава, где отдают многочисленные анастомозы к производным ствола плечевого сплетения. Дистальнее, к образовавшейся структуре, присоединяется второй грудной спинномозговой нерв. На правой конечности два краниальных (последние шейные) спинномозговых нерва объединяются между собой сразу после выхода из межпозвоночных отверстий, образуя единый ствол плечевого сплетения, направляющийся к свободной конечности. Затем к нему присоединяются первый и второй грудные спинномозговые нервы, в непосредственной близости к задней поверхности капсулы плечевого сустава. В результате, плечевое сплетение до разделения на нервы, иннервирующие мышцы-сгибатели и разгибатели, представлено единым общим стволом, содержащим в себе волокна всех четырех спинномозговых нервов (рисунок 1В). Таким образом, у земноводных и пресмыкающихся нерв сгибателей дает начало срединному и локтевому нервам, а нерв разгибателей гомологичен лучевому нерву. По данным М. Л. Голубева (1992) и М. В. Веселовского (1993), у земноводных и пресмыкающихся мышечно-кожного нерва не было отмечено. Данные о плечевом сплетении у птиц великолепно изложены в работах Г. Д. Канделаки (1983), который подчеркивал, что: «Сочетания спинномозговых нервов, образующих плечевое сплетение птиц, значительно отличаются не только у представителей разных видов, но и у отдельных особей одного и того же вида». В подавляющем большинстве случаев, постоянными участниками формирования плечевого сплетения являются два каудальных (шейных) и один краниальный (грудной) спинномозговые нервы, к которым могут присоединиться один или два переднележащих шейных нерва и один заднележащий грудной нерв или по одному передне- и заднележащему нерву. У голубя плечевое сплетение формируется из пяти спинномозговых нервов (сегментный состав – С13,С14,С15, Th1, Th2) [8,13]. При этом каудальный и краниальный стволы образуются соединением соответствующих пар спинномозговых нервов, а средний ствол, располагающийся между ними, представлен одним спинномозговым нервом. Эти стволы делятся на дорсальную и вентральную ветви (рисунок 1Г). Плечевое сплетение у курицы формируется волокнами четырех спинномозговых нервов (сегментный состав – С14, С15, Th1, Th2). Два краниальных нерва сплетения объединяются в краниальный, а два каудальных – в каудальный ствол сплетения [1,5]. Далее эти стволы делятся на вентральные и дорсальные ветви сплетения (рисунок 1Д). Данные о плечевом сплетении у млекопитающих представлены в работах А. Н. Микитюка (1983), В. В. Бобина (1986), Ю. Ю. Ильиной (1993), Е. К. Бондыревой (2003); А. И. Гирфанова (2009); Х. Б. Баймишев (2011). Еще в начале 19 века, А. Н. Северцовым (1908) в монографии, посвященной исследованию пятипалой конечности позвоночных, было выявлено, что при развитии плечевого сплетения в филогенезе, некоторые шейные сегменты редуцировались, а верхние грудные вошли в его состав. Общая схема образования плечевого сплетения у млекопитающих представлена на рисунке 1Е. По мнению Ю. Ю. Ильиной (1993), у млекопитающих, представителей отряда насекомоядных – ежа, плечевое сплетение образуется вентральными ветвями С5 – С8, Th1, Th2 (в 95 % случаев). В формировании плечевого сплетения участвуют первичные нервные стволы и только иногда у отдельных представителей обнаруживаются вторичные стволы, появление которых свидетельствует об увеличении количества сегментов, иннервирующих мышцы и, как следствие – расширение их функциональных возможностей [3,6,7]. Срединный нерв формируется из двух корешков, происходящих из всех вентральных ветвей, образующих сплетение. Мышечно-кожный нерв обособляется из сплетения проксимально и проходит через клюво-плечевую мышцу. У представителей отряда грызунов, по А. А. Ткачеву (1983), плечевое сплетение образуется вентральными ветвями (С5-С8, Th1), у некоторых животных (заяц, крыса, бобр) в сплетение входит пучок из Th2. У представителей отряда грызунов можно отметить различный уровень формирования из сплетения длинных стволов (мышечно-кожный, срединный и локтевой), идущих в сгибательной поверхности конечности. Изучением особенностей формирования плечевого сплетения у представителей отряда хищных (лисица, собака, кошка) занимались В. С. Абашидзе (1961), Е. М. Кулешова (1961), Д. Л. Асатиани (1970), которые пришли к выводу о том, что плечевое сплетение образуется вентральными ветвями С6 - С8, Th1. Кроме того, у собаки и лисицы найдена связь с Th2, а у кошки и С5 принимает участие в формировании сплетения. В. С. Абашидзе отмечал, что у собаки плечевое сплетение может перемещаться как в краниальном (С5 - С6), так и в каудальном (Th1 - Th2) направлениях. Уровень формирования нервных стволов у кошки происходит на плече более проксимально, чем у собаки. Таким образом, у хищных мышечно-кожный, срединный и локтевой нервы дифференцируются в проксимальной и средней трети плеча. У представителей отряда парнокопытных произошло уменьшение числа функций конечности в связи со специализацией – приспособление к бегу, что естественно отразилось на формировании уровня плечевого сплетения, которое образуется вентральными ветвями С6 - С8, Th1 и частично Th2. Так же, у парнокопытных отмечается значительное развитие коротких ветвей сплетения, идущих к хорошо развитым мышцам плечевого пояса. Таким образом, плечевое сплетение, представляющее собой очень крупное анатомическое образование, естественной границей которого, с одной стороны, является спинной мозг, а с другой, условной – подмышечная впадина, в основе своего строения имеет «принцип дихотомии». В формировании длинных ветвей плечевого сплетения у человека важное значение имеет участие сегмента C4 и Th2, а также изменчивость в формировании стволов из передней ветви С7. У земноводных вентральный нервный ствол, формируясь из плечевого сплетения, в локтевой области делится на срединную и локтевую ветви. У пресмыкающихся разделение вентрального ствола на эти нервы происходит более проксимально на плече, чем у земноводных. У ряда представителей этого класса прослежены различные формы обособления локтевого нерва (из вентрального нервного ствола). У млекопитающих выделяется ствол мышечно-кожного нерва. Срединный, локтевой и мышечно-кожный нервы произошли в филогенезе из одного источника – вентрального нервного ствола («нерва – сгибателей» низших позвоночных). Лучевой нерв произошел также из одного источника дорзального нервного ствола («нерва – разгибателей» низших позвоночных), который имеется в виде отдельного нерва у земноводных, пресмыкающихся и птиц. У млекопитающих лучевой нерв, являющийся филогенетически «древним» образованием, более дифференцирован, чем отдельные нервы сгибательной группы мышц конечности. Резюмируя ранее изложенное, нельзя не согласиться с высказыванием великого ученого морфолога А. Н. Северцова (1908) о том, что: «Плечевое сплетение – это своеобразный след эволюционных перестроек мышечной системы конечности, направленных на реализацию ее статолокомоторной функции. При общем уменьшении числа спинномозговых корешков, формирующих сплетение, наблюдаются усложнения в виде более интенсивных переплетений, появление связи между нервами, т.е. увеличивается степень их интеграции».
Библиографическая ссылка
Затолокина М.А., Мантулина Л.А. МОРФОФУНКЦИОНАЛЬНЫЕ ПАРАЛЛЕЛИ В РАЗВИТИИ ПЕРИФЕРИЧЕСКИХ НЕРВОВ ПЛЕЧЕВОГО СПЛЕТЕНИЯ В ЭВОЛЮЦИОННОМ АСПЕКТЕ // Современные проблемы науки и образования. 2016. № 2. ;URL: https://science-education.ru/ru/article/view?id=24200 (дата обращения: 18.07.2026).