



Видоспецифичность и устойчивость многих ритмов, а также возможность описания набора разных периодических составляющих активности и численности у различных видов животных привели к предположению о том, что спектр таких ритмов в определенных условиях может быть использован как таксономический признак [3]. При изучении многолетней популяционной динамики разных видов лесных насекомых-филлофагов также выявлялась определенная видоспецифичность у спектров ритмов популяционной динамики [8-10], отмечают ее и у других групп животных [1].
Учитывая это, целью настоящей работы было изучение и идентификация возможных таксономических различий в спектральных параметрах динамики очагов у насекомых-филлофагов из разных систематических групп. Для этого решались следующие задачи: - сравнительный анализ многолетней динамики площади очагов по статистическим характеристикам; -спектральный анализ многолетней динамики очагов массового размножения четырех видов лесных насекомых-филлофагов, сравнительный анализ их спектральных параметров и выявление возможных таксономических различий.
Материал и методика
Объектом исследования были временные ряды многолетней динамики вспышек массового размножения соснового шелкопряда (DendrolimuspiniL.), шелкопряда-монашенки (LymantriamonaсhaL.), обыкновенного соснового пилильщика (Diprion pini L.) и рыжего соснового пилильщика (NeodiprionsertiferGeoffr.) в лесах России. В работе использовались данные ФГУ Российского центра защиты леса [15].
Временные ряды популяционной динамики вспышек исследовались на наличие скрытых гармонических составляющих. Для каждого были построены спектры ритмов, а также рассчитаны периоды и мощности гармонических составляющих каждого такого спектра. При проведении исследований использовались программы спектрального анализа, находящиеся в собственности ИСиЭЖ СО РАН. Оценка спектральной плотности мощности проводилась методом Уэлча (Welch) [14]. Все расчёты выполнены с использованием свободной системы для математических вычислений GNU Octave [16,17]. В частности для оценивания СПМ методом Уэлча (Welch) использовалась функция pwelch из пакета расширений Octave-Forge [18]. Разработанный интерфейс позволил проводить обмен данными и всю обработку, включая выбор размера и количества окон, графическое представление данных и т.п. в диалоговом режиме. Для статистической обработки использован пакет программ PAST.
Результаты и обсуждение
Статистические оценки
Оценивая статистические средние значения динамики площади очагов, у 4 видов лесных насекомых-филлофагов выявлено, что максимальная площадь их за этот период наблюдалась у рыжего соснового пилильщика, а минимальная у соснового шелкопряда (табл. 1). Как показали результаты, у трех остальных видов насекомых-филлофагов средняя площадь очагов одного порядка (табл. 1). Кроме того, у всех четырех видов сходен и размах варьирования в динамике изменений площади очагов.
Таблица 1
Статистические параметры средней площади очагов массового размножения насекомых-филлофагов в лесах Российской Федерации в эруптивной фазе (1977–2007 гг.)
|
№ п/п |
Наименование вида насекомых-филлофагов |
Срок наблюдений, лет |
Средняя пло-щадь очагов, (М±m), га |
Среднее квадратич- ное откло- нение (σ) |
|
1 |
Сосновый шелкопряд |
29 |
9301,4±1148,9 |
6187,267 |
|
2 |
Шелкопряд- монашенка |
29 |
36340,9±4334,5 |
23341,74 |
|
3 |
Обыкновенный сосновый пилильщик |
29 |
24970,4±2019,2 |
10873,77 |
|
4 |
Рыжий сосновый пилильщик |
29 |
52580,7±5320,3 |
28650,79 |
Достоверность различий средних (с помощью t-критерия Стьюдента) оказалась высокой. Многолетние средние значения площади очагов у шелкопряда-монашенки с обоими видами пилильщиков достоверно различались на 5 % уровне значимости (t=2,4; P=0,05). Многолетняя средняя площади очагов соснового шелкопряда с обоими пилильщиками и пилильщиков между собой в эруптивной фазе достоверно различаются на 1 % уровне (t>4;P=0,01).
При оценке сходства колебаний площадей очагов у этих филлофагов мы обнаружили единственную значимую корреляцию между динамикой очагов соснового шелкопряда и обыкновенного соснового пилильщика (r=-0,403, P=0,05). Отрицательная корреляция может свидетельствовать о том, что многолетняя динамика вспышек массового размножения этих видов в наибольшей степени асинхронна. Мы предполагаем, что это может быть следствием проявления механизма разделения этих популяций. Несовпадение эруптивных фаз вспышек массового размножения у этих видов насекомых-филлофагов во времени может способствовать снижению уровня конкуренции за кормовое растение.
Сравнение многолетней динамики площади очагов
Графический анализ динамики площади очагов позволяет получить информацию не только о колебаниях их на территориях, но и выяснить привязку максимальных значений численности к определенным годам с их климатическими особенностями. В частности, можно визуально оценить совпадения максимумов у разных видов во времени.
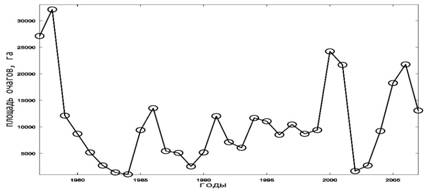
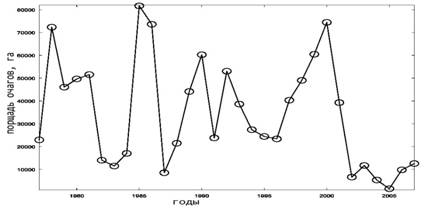
Сравнивая динамику очагов у соснового шелкопряда и шелкопряда-монашенки (рис. 1.1,1.2), заметна некоторая синхронность эруптивных фаз вспышек массового размножения. Так, в 1978, 1986 и в 2000 годах у этих видов была значительная численность, тогда как в 1983, 1996 и в 2002 годах прослеживается фазы минимума площади очагов.
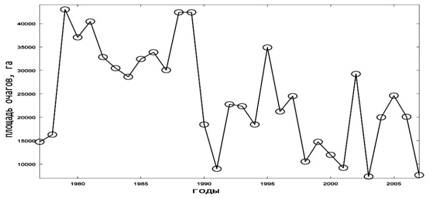
Сравнительный анализ динамики площади очагов у сосновых пилильщиков позволяет предполагать более тесную конкуренцию между этими видами, так как их эруптивные фазы не совпадают во времени. Когда один из видов находится в эруптивной фазе, у второго обычно происходит либо спад численности, либо даже ее депрессия. Исключение представляют два значительных подъема численности (1995 и 1997 гг.) и два небольших, происходившие сразу уобоих видов (рис.1.3, 1.4).
 1
1
 2
2
 3
3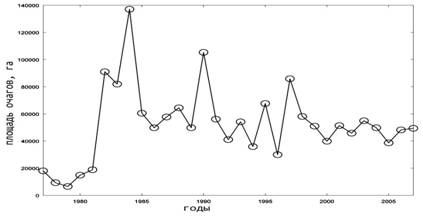 4
4
Рис. 1. Динамика площади очагов массового размножения насекомых-филлофагов в лесах Российской Федерации [15]. 1.Сосновый шелкопряд; 2. Шелкопряд-монашенка;
3. Обыкновенный сосновый пилильщик; 4. Рыжий сосновый пилильщик
Изменения площади очагов, расположенные на шкале времени (хронограммы), дают возможность визуально оценить цикличность у разных видовых популяций, но они не могут быть основой для точного сравнения популяционных циклов у разных видов. Более наглядно будет отображена цикличность, если представить материалы по динамике численности на частотной шкале, правда, при этом потеряется информация о самой динамике численности.
Анализ цикличности
Для этого представим материалы не в виде хронограмм, а используем анализ временных рядов и преобразуем их в частотные спектры (рис. 2).
Как показали результаты, для каждого спектра характерно небольшое число хорошо выраженных циклов. Более всего их имеют очаги рыжего соснового пилильщика (рис. 2, 4). Динамика всех рассматриваемых видов имеет циклы в близких полосах частот, особенно это касается высокочастотной части спектра. Близкие циклы наблюдаются чаще у разных систематических групп. У чешуекрылых цикличности часто совпадают с перепончатокрылыми (пилильщиками). Внутри одной группы такие близкие по значению ритмы фиксируются только в 2–3-летней полосе частот. Кроме того, у всех наблюдаемых насекомых отмечены ритмы динамики площади очагов в 6–8-летней полосе.
Еще одна особенность видовых спектров динамики вспышек состоит в том, что даже при одинаковых значениях колебаний их мощности могут значительно различаться. Так, у рыжего соснового пилильщика на спектре ритм 2-летний является субдоминантным по мощности, тогда как у обыкновенного соснового пилильщика этот ритм хотя и имеется, но мощность его незначительна (рис. 2, 3,4).
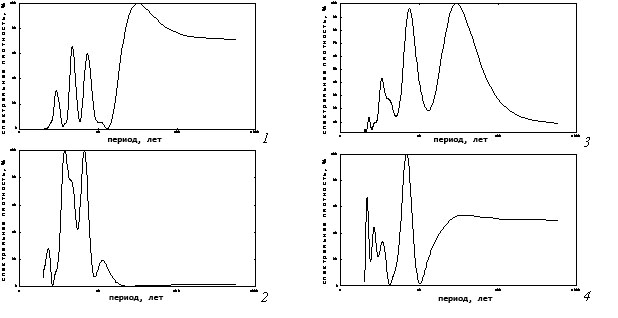
Рис. 2. Частотные спектры многолетней динамики площади очагов массового размножения насекомых-филлофагов (1 – Сосновый шелкопряд; 2 – Шелкопряд-монашенка;
3 – Обыкновенный сосновый пилильщик; 4 – Рыжий сосновый пилильщик)
Заметные различия в спектрах наблюдаются в 10–12-летней полосе частот. Здесь у шелкопрядов всегда имеются выраженные циклы, а у пилильщиков таковых не отмечается (табл. 2). Что касается низкочастотных периодических составляющих, то они специфичны для каждого вида (табл. 2).
Для соснового шелкопряда мы не располагаем данными о многолетней цикличности очагов массового размножения, но имеются сведения по динамике вспышек массового размножения «Европейской» географической популяции этого фитофага (163 года) [2]. Преобразование этого 163-летнего ряда данных в спектр ритмов позволило выделить наиболее мощные многолетние цикличности, свойственные динамике этого вида в Европе. Практически для всех полос частот, в которых мы нашли циклы численности этого вида, и в европейском спектре ритмов имеются соответствующие циклы. Причем, наиболее мощный цикл вспышек «Европейской» популяции соснового шелкопряда (12,1-летний) локализуется в той же полосе частот, что и в лесах Российской Федерации. Субдоминантный по мощности пик цикличности для «Европейской» популяции (6,9-летний) также совпадает по полосе частот с субдоминантным циклом, найденным для этого фитофага в лесах России (7,2-летний) (табл. 2). Близкими по частоте оказываются и многолетние циклы. Так, заметный цикл динамики (24,3-летний) выявлен нами на спектре ритмов «Европейской» популяции соснового шелкопряда. Что касается высокочастотных ритмов динамики вспышек, то тут у российских и европейских популяций наблюдается полное совпадение цикличности.
Для того чтобы оценить уровень сходства гармонических составляющих многолетних ритмов динамики площади очагов у шелкопряда-монашенки, мы также сравнили данные, полученные для этого вида по очень длинному ряду наблюдений (157 лет), опубликованные Е.Н Белецким [2]. Для «Европейской» географической популяции шелкопряда-монашенки, в частности, характерны следующие гармонические составляющие: 11,8, 6,8, 3,9 и 2,2-летние [9].
К настоящему времени нам не удалось в литературе обнаружить каких-либо данных по спектральному анализу популяционной динамики обыкновенного соснового пилильщика и рыжего соснового пилильщика. У обыкновенного соснового пилильщика за изученный период были идентифицированы четыре основных ритма. Самыми мощными из них были: 29,4- и 7,4 летний ритмы (табл. 2).
Таблица 2
Соотношение периодов и мощностей циклов в динамике площади очагов насекомых-филлофагов в лесах Российской Федерации
|
№ |
Период, лет Вид |
35-40 |
25-30 |
10-12 |
6-8 |
4-5 |
3-3,9 |
2,5-2.9 |
2-2,4 |
|
1. |
Сосновый шелкопряд |
|
31.1 165 |
10.5 9,3 |
7.2 100 |
4.6 108 |
|
2.9 50 |
|
|
2 |
Шелкопряд-монашенка |
|
|
11.4 447 |
6.7 2294 |
|
3.7 2313 |
|
2.3 642 |
|
3 |
Обыкновенный сосновый пилильщик |
|
29.4 694 |
|
7.4 670 |
|
3.3 298 |
|
2.2 91 |
|
4 |
Рыжий сосновый пилильщик |
37.3 1979 |
|
|
6.9 3685 |
|
3.4 1240 |
2.6 1646 |
2.1 2481 |
Примечание: верхняя цифра, период, нижняя – мощность (ед. спектр. плотн.)
Два высокочастотных ритма были значительно менее мощными. Как показали результаты, у рыжего соснового пилильщика, в отличие от предыдущего вида, доминировал среднечастотный ритм (6,9 летний), вторым по мощности был самый низкочастотный ритм (37,3 летний). Еще одним заметным отличием от предыдущего вида пилильщика было то, что серия высокочастотных ритмов имела достаточно высокую мощность. Сравнительный анализ полученных нами спектральных параметров этих двух видов насекомых-филлофагов показал, что по большинству средних и высоких частот гармоники у них близки. Различия имеются только в низкочастотной части спектра. Цикличность вспышек рыжего пилильщика на десятилетие медленнее, чем самый низкочастотный ритм у обыкновенного соснового пилильщика (табл. 2). В остальном же многолетняя цикличность у пилильщиков сходная, и в средних и высоких частотах они имеют близкие по значению ритмы. Как уже отмечено выше, заметным отличием были различия по мощности ритмов (табл. 2).
Полученные результаты позволяют предполагать, что из всех 4 рассматриваемых нами видов лесных насекомых-филлофагов наибольший биотический (вспышечный) потенциал характерен для рыжего соснового пилильщика, а наименьший – для соснового шелкопряда. Мы предполагаем, что это обусловлено высокой экологической пластичностью рыжего соснового пилильщика, холодостойкостью [5], что детерминирует его широкое распространение в северных широтах и, соответственно, расширение ареала. Высокая экологическая пластичность обеспечивает высокую скорость и интенсивность отклика на фактор абиотического стресса (весенне-летние засухи) и, соответственно, высокую скорость адаптации популяции к благоприятным изменениям биохимического состава кормового субстрата (хвои). Определенную позитивную роль в этом может выполнять и заметная приуроченность пространственной структуры очагов массового размножения рыжего соснового пилильщика к ослабленным насаждениям [5]. Ранее нами показано, что интенсивность реакции ослабленных древостоев на абиотический стресс (засуху), имеющих потенциально более низкую энтоморезистентность, заметно выше, чем древостоев за границами очагов [7,19].
Выводы
Как показали результаты, у насекомых-филлофагов из разных систематических групп выявлены таксономические различия в колебаниях площади очагов. По статистическим характеристикам динамика площади очагов у этих видов насекомых хорошо различается по средним многолетним значениям. Наибольшая средняя многолетняя площадь очагов в эруптивной фазе вспышек массового размножения выявлена у рыжего соснового пилильщика. Субдоминантом по площади очагов является шелкопряд-монашенка, затем обыкновенный сосновый пилильщик, и последнее место занимает сосновый шелкопряд. Близкого сходства в динамике вспышек у этих видов не выявлено, но характер динамики вспышек массового размножения соснового шелкопряда и обыкновенного соснового пилильщика продемонстрировали значимую отрицательную корреляцию.
Для всех изученных нами видов построены частотные спектры многолетней динамики вспышек массового размножения. Для каждого из них характерно небольшое количество гармонических составляющих. Все спектры близки по высокочастотным ритмам многолетней динамики очагов. Остальные колебания у них не совпадают по полосам частот.
Периодические составляющие колебаний площади очагов, даже если они у различных видов близки по значению, хорошо различаются по мощности.
Каждый из рассмотренных видов имеет специфичный спектр многолетних колебаний площади очагов массового размножения. Можно обозначить такой спектр, как видоспецифичный. Различия в спектрах внутри таксономических групп заметно меньше, чем между группами, то есть рассмотренные спектры ритмов динамики очагов массового размножения различаются таксономически.
Рецензенты:Усольцев В.А., д.с.-х.н., профессор, профессор Уральского государственного лесотехнического университета, г. Екатеринбург;
Петрова И.В., д.б.н., заведующий лабораторией популяционной биологии растений и динамики леса ФГБУН «Ботанический сад Российской академии наук», г. Екатеринбург.
Библиографическая ссылка
Колтунов Е.В., Ердаков Л.Н. ТАКСОНОМИЧЕСКИЕ ОСОБЕННОСТИ ЦИКЛИЧНОСТИ ДИНАМИКИ ОЧАГОВ МАССОВОГО РАЗМНОЖЕНИЯ ЛЕСНЫХ НАСЕКОМЫХ-ФИЛЛОФАГОВ В ЛЕСАХ РОССИИ // Современные проблемы науки и образования. 2015. № 2-1. ;URL: https://science-education.ru/ru/article/view?id=17524 (дата обращения: 07.07.2025).