



Процесс апоптоза является запрограммированной гибелью клеток, который может происходить при нормально протекающих условиях окружающей среды для организма, и может носить не всегда положительный характер. Исследования этого процесса за последние годы расширили представления о нем как о запрограммированной гибели клеток. Нас интересовали исследования этого процесса в области неврологии. Наиболее часто встречаются данные по изучению ишемии головного мозга, связанной с нарушением кровоснабжения тканей, данные повреждения запускают программы самоуничтожения клеток [2].
В ранних исследовательских работах гибель нейронов при острой церебральной ишемии интерпретирована как некроз ткани мозга. На современном этапе развития нейронауки существуют данные о том, что признаки апоптоза наблюдаются во всех структурах нейро-глиальной популяции, преимущественно повреждаются олигодендроциты. При этом количество погибших клеток возрастает в зависимости от продолжительности ишемии мозга. Существуют данные о том, что число апоптозных клеток максимально увеличивается на более длительном сроке воздействия экзогенным фактором. Эти данные свидетельствует о том, что реанимационные мероприятия, проводимые на ранних сроках воздействия экзогенным фактором, могут защитить поврежденную ткань мозга [4]. Современные исследования травматического повреждения спинного мозга рассматривают два основных взаимосвязанных механизма гибели клеток: апоптоз и некроз. Оба типа клеточной смерти имеют место быть в травмированном спинном мозге, при этом апоптоз рассматривается как один из путей клеточного обмена. На ранних этапах повреждения тканей спинного мозга наблюдается некроз – конденсация хроматина в размытые массы и деградация органоидов, позже происходит разрушение мембран, и только после этого запускается апоптоз, необходимый для обновления клеточного пула нервной ткани, её клеточной дифференцировки и развития. Как правило, апоптоз развивается при действии менее сильного повреждающего фактора, запуская внутренние энергозависимые механизмы самоуничтожения клетки – наблюдается конденсация ядерного хроматина, сморщивание тела клетки при сохранной цитоплазматической мембране, затем происходит разделение на отдельные фрагменты цитоплазмы, содержащие хроматин [5; 6]. Апоптоз в спинном мозге происходит в следующей последовательности – гибель нейронов, микроглии, олигодендроглии, приводя к общей дегенерации. Таким образом, апоптоз нейронов приводит к существенной потере числа активных клеток, растущей в прогрессии, а апоптоз глии препятствует выживанию и прорастанию оставшихся волокон, что выражается в отсутствии полноценной регенерации в спинном мозге. Изучение апоптоза при травматическом повреждении спинного мозга является очень перспективным с точки зрения возможности влияния на патологический процесс [5; 6].
Наряду с процессами гибели клетки изучались механизмы восстановления нервных клеток, одним из которых является нейрогенез. Процессы апоптоза и нейрогенеза – два взаимодополняющих механизма, направленных на поддержание гомеостаза в организме. Нейрогенез – процесс образования новых нервных клеток, в котором важную роль играет транскрипционный фактор Pax6 – регулятор пролиферации, дифференцировки и созревания нейронов, а также глиогенеза (процесс образования новых глиоцитов). В научной литературе приводятся факты об экспрессии Pax6 в стволовых клетках-предшественниках (радиальная глия), а также в коре и среднем отделе головного мозга. Экспериментально доказано, что большинство Рах6-позитивных клеток сохраняется и во взрослом мозге, но интенсивность нейрогенеза становится менее выраженной [3; 7]. Существуют исследования, в которых показано, что у крыс Pax6 экспрессируется в пролиферирующих нейрональных клетках непосредственно перед началом нейрогенеза. Данные клетки выявлены в переднем, заднем отделах головного мозга и спинном мозге. Полученные данные подтверждают функциональную активность Pax6 в нейронах животных, которые играют важную роль в пролиферации и в поддержании прогениторных клеток, нейрональной дифференцировке, в антиапоптозной функции [3; 5].
Цель исследования: изучить процессы апоптоза и нейрогенеза в нейро-глиальных популяциях спинного мозга молодых крыс после экзогенного воздействия.
Материалы и методы исследования
В нашем исследовании мы оценивали губительное влияние алкоголя и табачного дыма на нейроны спинного мозга животных. Эксперимент проводился на крысах пубертатного периода: одной группе животных (N=30) с помощью зонда внутрижелудочно вводился алкоголь (пиво «Балтика» 8%), другой группе (N=30) создавали с помощью специальной камеры условия пассивного курения (использовались легкие сигареты фирмы Winston). В зависимости от вида воздействия животные разделены на группы по 6 крыс согласно длительности воздействия (1, 3, 6, 12 часов) – экспериментальные группы. По окончании срока воздействия производился забор и фиксация материала (спинной мозг, шейно-грудной и поясничный отделы). Гистологические препараты были приготовлены по классической гистологической методике, окрашены тионином по методу Ниссля в модификации И.В. Викторова для определения степени хроматофилии нервных клеток; для выявления и оценки процессов нейрогенеза и апоптоза использовали иммуногистохимические методы и метод TUNEL.
На гистологических микропрепаратах проводился подсчет количества клеток, вступивших в процесс апоптоза, на разных сроках воздействия. Установление апоптоза в нейронах осуществлялось в результате окрашивания в клетках разрывов в цепи ДНК методом TUNEL. Для того чтобы иметь наиболее полную картину о процессах деградации и регенерации в нейронных популяциях спинного мозга, проводили параллели между количеством клеток, вступивших в апоптоз, и числом клеток с деструктивными изменениями, которые оценивались по степени хроматофилии их цитоплазмы [1; 3]. Хромофильное вещество – это вещество Ниссля, т.е. скопления цистерн в комплексе Гольджи с продуктами обмена клетки. Можно предположить, что деструктивные процессы идут не только в хроматине клеток, демонстрируя процесс апоптоза, но и в других структурных компонентах клетки (степень хроматофилии), приводя в общей массе к деструктивным и, возможно, необратимым процессам – к гибели клеток. В своих исследованиях мы сравнили количество клеток, вступивших в процесс апоптоза и пришедших в состояние деструкции (по степени хроматофилии – это тотально-гиперхромные, сморщенные клетки и клетки-тени) [4]. Параллельно оценивали процессы нейрогенеза, изучая синтез фактора Рах6 нейрональными клетками моторных ядер спинного мозга. Фактор транскрипции Рах6 (зеленые светящиеся точки) является ранним маркером нейрогенеза в нервных клетках. Статистическую обработку полученных результатов проводили с использованием пакета программ STATISTICA 6.0. Для выбора критерия оценки значимости различий проверяли соответствие формы нормального распределения, используя критерий χ2, а также равенство генеральных дисперсий с помощью F-критерия Фишера. Общее межгрупповое различие оценивалось при помощи Н-критерия Крускала-Уоллиса. В случае обнаружения различия нескольких выборок проводили множественное сравнение с использованием непараметрических вариантов критериев Даннета (для сравнения всех выборок с контрольной выборкой) и Ньюмена-Кейлса. Для сравнения двух опытных выборок (определенной популяции на определенный срок) использовали U-критерий Манна-Уитни.
Результаты исследования и их обсуждение
Результаты исследования показали, что после воздействия алкоголем клетки, вступившие в апоптоз, выявлены у животных контрольной и экспериментальных групп (табл. 1). Анализ данных позволил выявить, что количество апоптозных клеток после воздействия алкоголем значительно уменьшается на сроке 6 часов, на сроке 1 час равно нулю, на сроке 3 часа выявлены самые высокие значения. После воздействия табачным дымом наименьшее число нейронов, выступивших в апоптоз на сроке 1 час, после 12 часов воздействия они отсутствуют, наибольшее их количество наблюдалось после трехчасового воздействия экзогенного фактора (рис. 1). В экспериментальных группах, в которых наблюдалось уменьшение числа апоптозных нейронов, увеличивалось число клеток с деструктивными изменениями (тотально-гиперхромные, сморщенные клетки, клетки-тени). Нейроны, вошедшие в состояние деструкции, уже не смогут выйти из него, т.е. включается процесс некроза [6; 7].
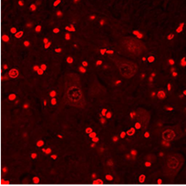 А
А 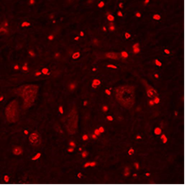 Б
Б 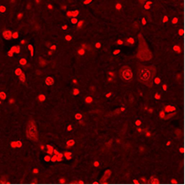 В
В
Рис. 1. Апоптоз в нейронах спинного мозга крыс: А – контрольная группа; Б – через 3 часа после воздействия алкоголем; В – через 3 часа после воздействия табачным дымом.
Как уже было выше сказано, наряду с процессами гибели клеток могут активизироваться процессы нейрогенеза, направленные на восстановление клеточного резерва тканей, что позволяет поддерживать гомеостаз [8; 9]. Для оценки процесса нейрогенеза подсчитывали количество клеток, синтезирующих транскрипционный фактор нейрогенеза Рах6. В результате оказалось, что данный процесс происходит в нервной ткани и в нормальных условиях существования организма.
Однако после воздействия алкоголем наблюдалось полное угнетение процессов нейрогенеза через 1 и 3 часа после воздействия, на сроке 6 часов его некоторая активизация и полное угнетение после 12 часов. После воздействия табачным дымом процессы нейрогенеза имеют другую характеристику. Через 1 час после воздействия они значительно активны, через 3 часа и в последующие сроки они подавлены, и не наблюдается их восстановления после 12 часов воздействия (рис. 2).
 А
А 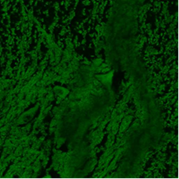 Б
Б  В
В
Рис. 2. Рах6-позитивные нейроны в спинном мозге крыс: А – в контрольной группе; Б – после воздействия алкоголем (6 часов); В – после воздействия табачным дымом (1 час).
Процессы апоптоза и нейрогенеза в нервных клетках моторных ядер спинного мозга после экзогенного воздействия
|
Сроки воздействия |
Алкоголь
|
Табачный дым |
||||||||
|
Клетки в состоянии деструкции, абс. число |
Клетки в состоянии апоптоза, абс. число |
Pax 6-позитивные клетки, абс. число |
Клетки в состоянии деструкции, абс. число |
Клетки в состоянии апоптоза, абс. число |
Pax 6-позитивные клетки, абс. число |
|||||
|
тотально-гиперхромные |
сморщенные |
клетки-тени |
тотально-гиперхромные |
сморщенные |
клетки-тени |
|||||
|
контроль |
0 |
0 |
0 |
34,5 [5,5; 44,5] |
2 [0; 3] |
0 |
0 |
0 |
34,5 [5,5; 44,5] |
2 [0; 3] |
|
1 час |
0 |
0 |
0 |
0 |
0 |
1* [0; 3] |
0 |
1,7* [0; 5] |
10,5* [1,5; 20,5] |
28,89* [5,13; 61,29] |
|
3 часа |
3,94* [0; 10] |
0 |
0 |
30* [4,5; 40] |
0 |
1,49* [0; 5] |
0 |
0 |
17,5* [3; 27,5] |
0 |
|
6 часов |
3,78* [0; 10] |
0 |
0 |
17,5* [6,5; 20] |
18,03* [5,26; 51,06] |
6,15* [0; 19,7] |
1,2* [0; 3] |
5,6* [0; 14,2] |
14* [2,5; 24] |
0 |
|
12 часов |
5,98* [0; 15] |
1,69* [0; 10] |
0 |
26* [4; 36] |
0 |
13,13* [11,3; 22,2] |
2,35* [1,9; 6,2] |
1,28* [0; 3,3] |
0 |
0 |
* – наличие статистически значимых различий между контролем и экспериментальными группами по критерию Манна-Уитни (при p<0,05).
Заключение
Проведя анализ полученных результатов, можно сделать вывод о том, что нейроны спинного мозга проявляют пластичность после воздействия табачным дымом на сроке 1 час и на сроке 6 часов после воздействия алкоголем, а также в контрольной группе животных. Это является очередным подтверждением включения компенсаторно-приспособительных возможностей нервных клеток спинного мозга при экзогенном воздействии. Также подтвердились уже имеющиеся данные в литературе о том, что с удлинением срока воздействия процессы запрограммированной гибели клеток и нейрогенеза угнетаются, возрастает число клеток с деструктивными признаками, что свидетельствует о присутствии некротических процессов. Проведенное исследование продемонстрировало, что табачный дым оказывает более губительное действие на спинной мозг экспериментальных животных в сравнении с действием алкоголя (легкий алкогольный напиток).
Рецензенты:
Мучкина Е.Я., д.б.н., проф. кафедры экологии и природопользования ИЭУиП ФГАОУ ВПО «СФУ», г. Красноярск;
Зайцева О.И., д.м.н., проф., старший научный сотрудник лаборатории этногенетических и метаболических проблем нормы и патологии, Институт медицинских проблем Севера СО РАМН, г. Красноярск.
Библиографическая ссылка
Устинова Т.И., Медведева Н.Н., Малиновская Н.А. ПРОЦЕССЫ АПОПТОЗА И НЕЙРОГЕНЕЗА В НЕЙРОНАХ МОТОРНЫХ ЯДЕР СПИННОГО МОЗГА МОЛОДЫХ КРЫС ПОСЛЕ ЭКЗОГЕННЫХ ВОЗДЕЙСТВИЙ // Современные проблемы науки и образования. 2015. № 2-1. ;URL: https://science-education.ru/ru/article/view?id=17185 (дата обращения: 13.07.2026).