



Данные о медленных отклонениях электрического потенциала коры мозга от постоянного уровня встречаются еще в работах, проведенных в последнее столетие [1-3]. В конце 80-х и начале 90-х годов прошлого столетия была подробнее описана сверхмедленная активность головного мозга и показано, что омега-потенциал может служить универсальным показателем, отражающим уровни бодрствования, стрессорную устойчивость, компенсаторно-приспособительные возможности организма.
Один из видов сверхмедленной электрической активности головного мозга - это устойчивый потенциал милливольтового диапазона – омега-потенциал [4; 5]. Колебания омега-потенциала отражают процессы, происходящие в головном мозге и его подкорковых образованиях. Следует отметить, что все структуры мозга, за исключением гипоталамических ядер, характеризуются одинаковым порядком частоты сверхмедленного ритма, а частота колебаний становится выше в особенности после их стимуляции относительно других образований мозга. Более высокую интенсивность частоты сверхмедленных колебаний потенциалов в гипоталамусе можно объяснить особенной чувствительностью этой области к действию многих высокоактивных веществ, таких как нейрогормоны, а именно серотонин, гистамин, норадреналин и пр. Омега-потенциал свидетельствует о мере координированности межсистемных и межорганных нейрогуморальных взаимоотношений.
Результаты исследований имеют не только теоретическое значение, но и могут найти широкое практическое применение в области медицины, гигиены, спорта, а также в интересах различных правоохранительных и особенно военных ведомств, так как исследование проблемы адаптации организма к гипоксии – это и изучение возможностей приспособления организма к экстремальным условиям, и повышение его общей резистентности.
Цель работы – охарактеризовать омега-потенциал у здоровых и больных лиц, его изменения после комплексного курортного лечения, включающего курс нормобарической интервальной гипоксической тренировки в санатории «Нальчик», а также выявить механизмы приспособления животных к недостатку кислорода.
Материал и методы исследования. Обследовано 55 пациентов, из них 25 здоровых отдыхающих и 30 страдающих неспецифическим хроническим обструктивным бронхитом, бронхиальной астмой. Фоновый омега-потенциал (ОП) в отведениях лоб-тенар кисти и вертекс- тенар кисти (F-T, V-T) регистрировался на приборе «Омега-4», с масштабом 1 mV, 0.3 s. Омегограмма записывалась до и после курортного лечения, включавшего для обследованных нормобарическую интервальную гипоксическую тренировку (ИГТ), курс которой состоял из 15 сеансов. В первых пяти сеансах содержание кислорода в подаваемой газовой смеси составляло 16%, во вторых пяти – 14%, в третьих – 12%.
Рабочий и вспомогательный электроды подключались к полярографической установке (рисунок 1), и снималась полная вольтамперная характеристика рабочего электрода (I- j- кривая) в физрастворе, уравновешенном с воздухом при нормальном барометрическом давлении (первый раствор), а затем в аналогичном растворе, предварительно лишенном кислорода добавлением щепотки сульфита натрия (0,1-1,0 г на 100 мл раствора - II раствор). Полученные в I и II растворе полярограммы позволяли судить о пригодности электрода для определения кислорода и выбрать рабочую точку на I- j-кривой в I растворе.
Рабочая точка на I- j-кривой в I растворе - значение потенциала поляризации, при котором в дальнейшем будет осуществляться восстановление О2 - для золота этот потенциал в пределах 0,50-1,20 В, выбиралась в самом начале плато диффузионного тока.
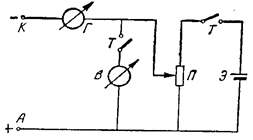
Рис. 1. Схема простейшей установки (схема Гейровского), использующейся для полярографического определения кислорода:
Э - гальванический элемент; Т - тумблер; П - проволочное переменное сопротивление 0,3-1,0 ком; В - высокоомный вольтметр; Г - чувствительный зеркальный гальванометр с набором шунтов;
К - клемма для подключения рабочего электрода (катода); А - клемма для включения вспомогательного электрода (анода)
При изучении процесса адаптации к гипоксии кроликов (n=13) было сформировано 2 группы: контрольная (нетренированные) и опытная (адаптированные). Приспособление подопытного поголовья к нехватке кислорода осуществлялось путем проведения ритмической гипоксической тренировки как в норме, так и в условиях гипоксии в барокамере. Исследования проводились в лабораторной барокамере, в которой создавалось разрежение воздуха.
Полученный цифровой материал обрабатывали методом вариационной статистики с использованием персонального компьютера на основе пакета программ анализа данных Statistica 6.0. Из статистических показателей использовали среднюю арифметическую (Х), ошибку средней арифметической (mх). Различия между сравниваемыми значениями устанавливали по критерию Стьюдента при трех уровнях вероятности: Р>0,95, Р>0,99 и Р>0,999.
Результаты исследования и их обсуждение. До интервальной гипоксической тренировки у большинства как здоровых, так и больных обследованных наблюдались: длительный период стабилизации фонового омега-потенциала, низкие значения омега-потенциала, большая разница между начальным значением омега-потенциала и его величиной при выходе на плато, что свидетельствовало о низком уровне бодрствования.
В результате проведенных обследований у всех лиц, как у здоровых, так и с неспецифическим хроническим обструктивным бронхитом, после комплексного лечения выявили резкое снижение продолжительности выхода исходных значений омега-потенциала на плато (таблицы 1, 2).
Таблица 1
Динамика показателей омега-потенциала у здоровых лиц в условиях нормоксии
|
Время стабилизации фонового ОП (min), M±mх |
Значение ОП при выходе на плато (mV), M±mх |
Начальное значение ОП (mV), M±mх |
Разность между начальным значением и величиной ОП при выходе на плато (min), M±mх |
||||
|
до ИГТ |
после ИГТ |
до ИГТ |
после ИГТ |
до ИГТ |
после ИГТ |
до ИГТ |
после ИГТ |
|
4,03± 0,04*** |
1,95± 0,02 |
-0,49 ±0,7 |
-7,88± 0,9*** |
-2,79± 1,6 |
-0,95± 1,0 |
-2,3± 1,4 |
-0,07± 0,04 |
Примечание: достоверно при *Р>0,95, *Р>0,99, *Р>0,999.
Установлено, что в результате приспособления к гипоксии в условиях поочередного нормоксического и гипоксического воздействия значения омега-потенциала после выхода на плато в целом были в тех же начальных пределах, или же имели место более высокие величины. При наличии колебаний омега-потенциала минутного диапазона до выхода на плато у неадаптированных людей, после курса интервальной гипоксической тренировки их не наблюдалось. Имеет место характерная тенденция в сторону сближения значений исходного омега-потенциала с его уровнем после выхода на плато.
Таблица 2
Динамика показателей омега-потенциала у лиц с неспецифическим хроническим обструктивным бронхитом в условиях нормоксии
|
Время стабилизации фонового ОП (min), M±mх |
Значение ОП при выходе на плато (mV), M±mх |
Начальное значение ОП (mV), M±mх |
Разность между начальным значением и величиной ОП при выходе на плато (min), M±mх |
||||
|
до ИГТ |
после ИГТ |
до ИГТ |
после ИГТ |
до ИГТ |
после ИГТ |
до ИГТ |
после ИГТ |
|
5,17± 0,06*** |
3,55± 0,04 |
-4,03± 0,05 |
-16,03± 1,3*** |
-30,18± 1,5 |
-30,18± 1,7 |
-26,15± 1,3*** |
-14,15± 0,9 |
Примечание: достоверно при *Р>0,95, *Р>0,99, *Р>0,999.
Как видно из представленных данных, изменения показателей фонового омега-потенциала, к которым привела интервальная гипоксическая тренировка у здоровых (таблица 1) и больных неспецифическим хроническим обструктивным бронхитом (таблица 2) людей, имеют сходную динамику, но при этом имеет место и ряд отличий. Так, у здоровых лиц время стабилизации омега-потенциала по сравнению с больными неспецифическим хроническим обструктивным бронхитом меньше и после адаптации к интервальной гипоксической тренировке уменьшается еще примерно в два раза, тогда как у больных адаптация к гипоксии приводит к уменьшению лишь примерно в полтора раза. Следует отметить и то, что устойчивое значение омега-потенциала у больных людей по сравнению со здоровыми было отчетливо меньше и соответствовало низкому уровню бодрствования.
Установленные особенности фонового омега-потенциала как у приспособленных к недостатку кислорода здоровых людей, так и у таковых с выраженным неспецифическим хроническим обструктивным бронхитом характеризуют повышение приспособительных качеств и неспецифических факторов реактивности организма к кислородному голоданию.
Совершенно отчетливо прослеживается процесс адаптации к гипоксии и при анализе электрокортикограммы кролика. Электроэнцефалография продолжает оставаться одним из доступных методов постоянного слежения за количеством нейронов в мозге. Характеризуя электроэнцефалограмму кролика в состоянии спокойного бодрствования, ученые [6; 7] выявили, что она содержит в основном частоты дельта- и тета-диапазонов.
В наших исследованиях подъем кроликов до высоты 7000-8000 м характеризовался доминированием тета-ритма электрокортикограммы, от высот 8000 до 11000 м – превалированием дельта-волн, а на высоту 11000-12000 м – уплощением биоэлектрической кривой и снижением амплитуды потенциалов свыше.
Не установлено достоверных различий между группами кроликов по степени их подъема до высоты 7000 м, что в целом сопровождалось снижением амплитуды электрокортикограммы. Вероятно, нарастающее превалирование амплитуды тета-ритма может являться причиной снижения электрокортикограммы. Дальнейший подъем на высоту 8000 м сопровождался более существенными различиями между сравниваемыми группами кроликов. Так, у неадаптированных к гипоксии кроликов реакция на гипоксические условия сопровождалась резким увеличением амплитуды электрокортикограммы, в то время как у тренированных особей подобной реакции не было, а, наоборот, имело место еще более высокое снижение амплитуды биоэлектрической активности коры. Как и ожидалось, у опытной группы кроликов на высоте 8500 м продолжилось снижение амплитуды электрокортикограммы, тогда как на высоте 9000 м имело место резкое увеличение амплитуды биоэлектрической активности.
Характеризуя электроэнцефалограмму кролика на высотах 8000 и 9000 м и более, отметим, что здесь имеет место проявление медленных высокоамплитудных волн, с чем, вероятно, связано резкое повышение амплитуды электрокортикограммы, наблюдающееся у обеих групп особей.
Причиной увеличения амплитуды регистрируемой активности является расширение площади коры, которая охвачена повышенной синхронизацией. Имеющие место гиперсинхронизированные колебания на обширной площади коры человека способствуют достоверному повышению амплитуды электроэнцефалограммы, которую можно рассматривать как результат сложения полей пирамидных нейронов коры. Для возникновения генеза электроэнцефалограммы необходима одинаковая ориентация диполей нейронов и наличие положительной корреляции в их градуальной электрической активности. Данная точка зрения находит свое подтверждение и в гистологических исследованиях строения коры, которые свидетельствуют, что около 70% всех нейронов коры составляют пирамидные нейроны, имеющие совершенно четкую вертикальную ориентацию в направлении, поперечном поверхности коры [8].
В наших исследованиях у контрольной группы кроликов увеличение амплитуды электрокортикограммы происходило с высоты 7000 м (Р≤ 0,001), тогда как у опытной группы повышение синхронизации биоэлектрической активности коры зафиксировано только с высоты 8500 м (Р≤ 0,001). Различия в 1500 м между подопытными группами кроликов характеризуют адаптацию к гипоксическим условиям.
Если напряжение кислорода в коре головного мозга, независимо от тренированности кроликов, при увеличении высоты постепенно уменьшилось, то при последующем спуске – обратно повысилось, практически до исходных значений. При прочих равных условиях у опытной группы кроликов уровень напряжения кислорода в коре, в условиях нормы (63,8±1,4), был выше на 7,3 мм рт. ст., чем у контрольных аналогов (56,5±1,1), благодаря чему даже на высоте 8000 м этот показатель у адаптированных к гипоксии кроликов (43,2±0,6) был выше на 3,3 мм рт. ст. (Р≤ 0,001), чем у неадаптированных (39,9±1,1) (рисунок 2).
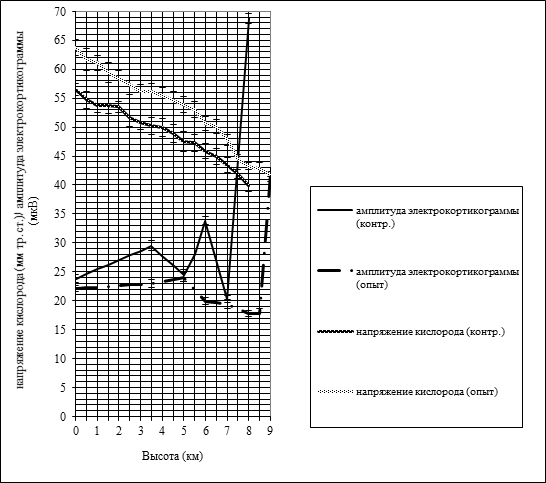
Рис. 2. Результаты изменения амплитуды электрокортикограммы и напряжения кислорода в коре головного мозга у адаптированных и неадаптированных к гипоксии кроликов
Выяснено, что у кроликов в результате возобновления мозгового кровотока рО2 восстанавливается со значительным овершутом, что свидетельствует о падении потребления кислорода мозговой тканью. Указанная тенденция может являться функциональной исходя из гипотезы о рефлекторном подавлении работы нейронов при гипоксии.
Считается, что в результате поднятия на высоты в организме происходит увеличение концентрации гемоглобина и эритроцитов в крови, которое осуществляется за счет выброса кровяных депо и сгущения крови, а затем – вследствие интенсификации кроветворения. Примерно через 10 суток быстрая фаза нарастания количества эритроцитов и содержания гемоглобина в крови завершается.
Таким образом, мы можем предположить, что при кратковременной адаптации к гипоксии главным механизмом, обеспечивающим повышающуюся потребность мозга в кислороде, является увеличение скорости мозгового кровотока, а при долговременной адаптации – увеличение плотности капиллярной сети мозга.
По результатам проведенных опытов можно констатировать высокую обусловленность изученных показателей – напряжения кислорода в коре головного мозга с биоэлектрической активностью. Эта зависимость, в результате подъема подопытных кроликов, независимо от их адаптированности, влияла на изменение, в частности на повышение амплитуды электрокортикограммы в результате достижения общего критического уровня напряжения кислорода.
Заключение. Таким образом, можно констатировать, что использование чередования нормоксических и гипоксических адаптаций в комплексном санаторно-курортном лечении - эффективный метод улучшения состояния здоровья и повышения работоспособности здоровых и больных хроническим обструктивным бронхитом людей, хороший метод профилактики.
Курс интервальной гипоксической тренировки в комплексном санаторно-курортном лечении приспосабливает организм к нехватке кислорода, увеличивает реактивную способность к ней, тем самым расширяя возможности фурье-стресс дифрактометра. Использование указанной адаптации организма к недостатку кислорода повышает приспособление организма, в частности антигипоксантную и антиоксидантную системы, и согласуется с исследованиями в этом направлении [9; 10].