



Исследование первичных спонтанных процессов лесообразования в долинах крупных рек позволяет понять ход, закономерности и механизмы эволюционно сложившегося цикла сукцессионных процессов, приводящих к формированию зональной лесной растительности. Для глубокого понимания этих процессов необходимо изучить закономерности изменения флористического состава и структуры первичных сообществ в ходе аллювиогенных сукцессий.
Цель исследования – изучение закономерностей изменения флористического состава и вертикальной, экоморфной структуры лесных и кустарниковых сообществ в ходе аллювиогенных сукцессий в долине среднего течения р. Лены.
Материал и методы исследования
Исследования проведены вдоль поперечной и продольной трансект на низкой, средней, высокой поймах, I, II надпойменных террасах Покровского, Якутского и Намского участков долины среднего течения р. Лены. Материалами послужили около 200 лесоводственно-геоботанических описаний, выполненных и проанализированных по традиционным методикам [7; 8], и созданная на их основе база данных в программе IBIS.
Участие видов и групп в фитоценозах определено методом расчета коэффициентов участия (КУ), предложенного В.С. Ипатовым [2]. Преимуществом использования этого коэффициента является возможность комплексно оценить встречаемость и обилие видов. Коэффициент участия рассчитан по формуле:
КУ=∑а х m / n² x 8, где n – число описаний, m – встречаемость группы видов, a – ранг группы, число 8 – максимальный ранг обилия группы. КУ может принимать значения от 0 до 1. Для анализа распределения древесных, кустарниковых пород и видов-доминантов травяного покрова построены топоклины изменения коэффициента участия основных видов вдоль градиента сукцессий. Латинские названия высших сосудистых видов растений приводятся по «Конспекту флоры Якутии» [3], мохообразных и лишайников – по сводке «Разнообразие растительного мира Якутии» [6].
Результаты исследования и их обсуждение
Интенсивная аллювиальность, переформирование речного русла, характерные для крупных меандрирующих рек, обеспечивают появление новых аллювиальных субстратов. Река Лена в своём среднем течении характеризуется большими объемами ежегодного аллювионакопления. По данным В.Л. Суходровского [9], А.В. Шестакова [10], у левого берега р. Лены за год отмечается намыв аллювия шириной до 10-12 (20) м. На вновь образованных прирусловых субстратах происходят масштабные сингенетические процессы. Сингенез начинается с заселения аллювиофильных ив, и через длительный, последовательный путь развития серия заканчивается на надпойменных террасах формированием коренных хвойных типов лесов.
Количество видов в сообществах, их обилие, экоценоморфный состав определяются внешними и внутренними факторами и колеблются на протяжении всего сукцессионного ряда. Существенные изменения видового состава происходят при повышении гипсометрического уровня поймы, что закономерно происходит с поднятием пойменных поверхностей в высоту в результате седиментации и снижения базиса эрозии.
Экогенетический ряд лесной растительности в пойме среднего течения р. Лены (рис. 1, 2) начинается с сингенеза растений на аллювиальных свежих песчаных и илисто-песчаных отложениях. Первыми поселяются аллювиофильные ивы: на отмытых песках – S. viminalis L., на илисто-песчаных – S. triandra L. и S. udensis Trautv. et C. A. Mey. С заселения аллювиофильных ив начинается I аллювиофильноивовая фаза развития лесной растительности поймы, включающая стадии: беспокровную (от 0 до 5 лет), хвощовую (от 5 до 15 (20) лет), крупнокорневищных гигромезофильных злаков (от 15 (20) до 30 (35) лет), мезо- и ксеромезофильных злаков и разнотравья (от 30 (35) до 40 (50) лет).
Сингенез травянистых растений на первых стадиях хаотичен, что обусловлено приносом зачатков водой, открытостью субстратов, отсутствием конкуренции. В первые годы наблюдаются прибрежно-водные (Alisma plantago-aquatica L.), однолетние и сорные виды. К 5-летнему возрасту большинство случайных видов элиминируется, в покрове повышается обилие Equisetum arvense L. Хвощовая стадия длится примерно до 10-15 лет до достижения ивовым древостоем среднего и приспевающего возраста. Затем увеличивается обилие гигрофильных корневищных злаков (Calamagrostis purpurea subsp. langsdorffii (Link) Tzvel., Bromopsis inermis (Leyss.) Holub, Agrostis gigantea Roth, Phalaroides arundinacea (L.) Rauschert), приспособленных к аллювионакоплению и поёмности. Способность выдерживать интенсивную аллювиальность возможна благодаря их свойству перемещать узел кущения вверх по мере отложения речного аллювия на поверхности почвы. Будучи сильными эдификаторами травяного покрова с мощными корневыми системами, высокими стеблями, широкими листовыми пластинками, эти злаки заглушают хвощ и начинают доминировать. Наряду со злаками появляются пойменные гигро- и гигромезофильные теневыносливые растения, также развивающие ползучие корневища: Naumburgia thyrsiflora (L.) Reichenb., Impatiens noli-tangere L., Ranunculus repens L. Степень покрытия почвы увеличивается до 90-95%, травостой смыкается, и ассоциация становится условно закрытой. Наступает вторая стадия динамики аллювиофильных пойменных ивняков – стадия крупнокорневищных гигро- и гигромезофильных злаков и соответствует возрастным группам приспевания-перестойности древостоя (от 15-20 до 30-35 лет). Сообщества этой стадии представляют ассоциации Salicetum viminalis bromopsidosum inermis, S. v. calamagrostidosum langsdorffii.
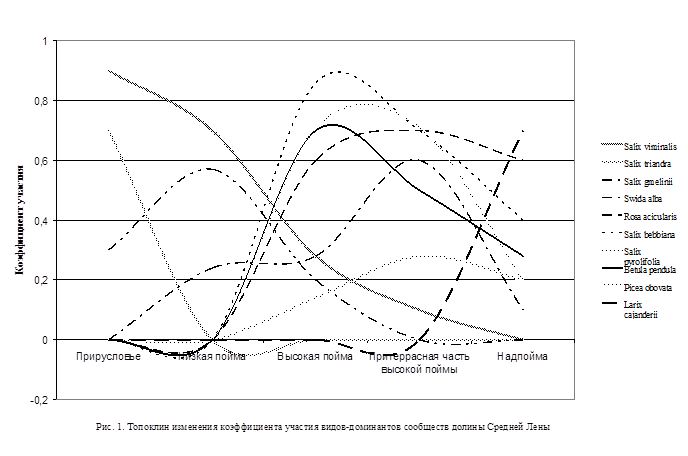
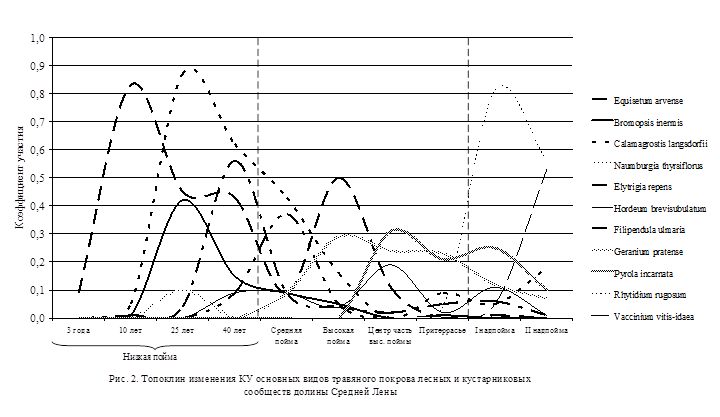
При переходе на уровень средней поймы в результате продолжающейся седиментации и понижения базиса эрозии наступает следующая стадия – ксеромезофильных злаков и высокотравья. Эта стадия занимает период примерно от 30 (35) до 40 (50) лет возраста древостоя. Представлена сообществами ассоциации Salicetum viminalis elytrigiosum repentis. Характерно резкое ослабление аллювиальности и поёмности. Аллювиофильноивовый древостой к этому времени достигает перестойного возраста. Усыхание крон, вывал деревьев постепенно приводят к повышению сквозистости крон, что способствует увеличению испарения с поверхности почвы. Изменение условий фитоценотической среды и почвенно-грунтовых условий обусловливает снижение участия гигро- и гигромезофильных злаков и крупнотравья, а в последующем – полную элиминацию наиболее влаголюбивых видов. Травяной покров начинает приобретать более мезофитный характер. Топоклин коэффициента участия видов травяного покрова иллюстрирует появление и постепенное увеличение роли Hordeum brevisubulatum (Trin.) Link, чаще преобладающего в этот период (рис. 2). Аллювиальные слаборазвитые почвы поймы постепенно развиваются в относительно плодородные аллювиальные дерновые. На этой стадии древостой из аллювиофильных ив почти полностью распадается, остаются единичные порослевые экземпляры.
Естественное возобновление аллювиофильноивовых сообществ под пологом древостоя отсутствует, поэтому эти ивняки существуют на низкой пойме лишь одно поколение древостоя. Сукцессионный ряд сообществ из аллювиофильных ив на средней пойме завершается, постепенно сменяясь на плоских элементах мезорельефа мезофильными разнотравно-злаковыми (с Hordeum brevisubulatum, Elytrigia repens (L.) Nevski, Geranium pratense L., Thalictrum simplex L., Galium boreale L. и др.), на выпуклых – остепненными лугами.
При выходе на уровень высокой поймы на дренированных межгривовых понижениях, на низких плоских увалах, ложбинах стока, по склонам стариц, грив, проток с более благоприятным гидротермическим режимом, на аллювиальных дерновых, дерново-глееватых почвах формируются ленточные сообщества из аллювиофобных видов ив с преобладанием Salix pyrolifolia Ledeb. или S. bebbiana Sarg. Они характеризуются преобладанием разнотравья в напочвенном покрове и развитием хорошо выраженного второго подъяруса из средних гигромезофильных (Swida alba (L.) Opiz, Spiraea salicifolia L.), мезофильных и мезотрофных кустарников (Rosa acicularis Lindl., Ribes glabellum (Trautv. et C.A. Mey) Hedl., R. pauciflorum Turcz. ex Pojark, Crataegus dahurica Koehne et Schneid.). Топоклин иллюстрирует повышение участия на высокой пойме Swida alba, Rosa acicularis (рис. 1). Травяной покров в основном представлен разнотравьем, опушечными, луговыми видами.
Большое суммарное водопотребление мезофитных ивовых сообществ способствует иссушению почвы. Постепенно комплекс условий, способствовавших разрастанию кустарниковых аллювиофобноивовых ассоциаций, значительно трансформируется. Увеличивается деградация травяного покрова, снижается сомкнутость подлеска. К этому времени понижается виталитет Salix bebbiana и S. pyrolifolia, наблюдается суховершинность, отмирание части крон, ослабление порослевого возобновления. Эти процессы приводят к снижению корневой конкуренции, освобождению экологических ниш, что благоприятствует появлению всходов и закреплению подроста других пород – в наших условиях Betula pendula Roth. В ненарушенных антропогенным воздействием фитоценозах дальнейшее развитие приводит к появлению в травяном покрове типичных лесных видов, например Pyrola asarifolia Michaux, P. rotundifolia L., Orthilia obtusata (Turcz.) Jutrz., изредка Vaccinium vitis-idaea L. (рис. 2). Они служат инициальными видами-индикаторами последующих смен – начала развития зональных типов леса. Также появляются эпифитный лишайник Xanthoria fallax (Hepp) Arnold и некоторые виды типичных мхов зональных коренных лесов, например Sanionia uncinata (Hedw.) Loeske. К этому времени факторы экотопа все более приближаются к условиям надпойменных террас, влияние гидрологического режима реки ослабевает. Постепенно образуется маломощная лесная подстилка и грубогумусовый перегнойный горизонт, формируются более плодородные дерновые перегнойные, дерново-лесные почвы.
С течением времени в центральной и притеррасной частях поймы на прибрежных склонах стариц, в межгривовых понижениях, на склонах грив аллювиофобноивовые сообщества сменяются березняками. Наступает березовая фаза развития лесной растительности поймы Средней Лены. Главной особенностью структуры березовых лесов из Betula pendula является более сложная вертикальная структура, проявляющаяся в 1-2-ярусности древостоя, наличии 2-подъярусного сильно развитого кустарникового подлеска, разреженного травяного покрова без ярусности и маломощной развитой лесной подстилки. Степень проективного покрытия травяного покрова довольно низка – в среднем 40%, варьирует от 30 до 80%. В верхнем ярусе подлеска господствует и высоконстантна Salix bebbiana, в нижнем – Rosa acicularis, Ribes glabellum. В травяном покрове постоянны Geranium pratense, Sanguisorba officinalis L., Galium boreale, Galium verum L., Hordeum brevisubulatum, Geranium pratense (95-100%-ная встречаемость). В сукцессионно более продвинутых лесах высоконстантна Pyrola asarifolia. Характерно отсутствие бореальных кустарничков. Среднее видовое богатство высокое – 34 вида. Напочвенный моховой покров отсутствует. Из лишайников редко наблюдаются отдельные пятна Peltigera aphtosa (L.) Willd., часто регистрируются эпифиты (Xanthoria fallax, Parmelia sulcata Taylor).
Через несколько поколений березового древостоя в спелых, перестойных насаждениях при условии наличия семенных деревьев в соседствующих фитоценозах появляются всходы ели. Фитомикроклимат березового древостоя благоприятно влияет на всходы ели посредством угнетения конкурирующих видов напочвенного покрова, смягчения резких перепадов температур в приземном слое воздуха и защиты от отрицательного влияния прямых солнечных лучей. Со времени проявления эдификаторной роли ели в насаждениях в сукцессионной серии наступает фаза еловых лесов (рис. 1). Ельники характеризуются сложным составом древостоя, 2-3-ярусностью, высокосомкнутостью (0,9-1,0). Подлесок из кустарников практически отсутствует, характерно наличие жизнеспособного подроста ели; травяной покров чаще деградирован (покрытие 10-15%). Высококонстантны Calamagrostis lapponica (Wahlenb.) C. Hurtm., Equisetum pratense Ehrh. и обычные «сквозные» виды – Thalictrum simplex, Sanguisorba officinalis, Iris setosa Pall. ex Link и др. Нередко с небольшим обилием встречаются виды лесного, лесоопушечного разнотравья: Valeriana capitata Pall.ex Link, Viola mauritii Tepl. Характерно сопровождение таёжного теневого мелкотравья: Trientalis europaea L., Adoxa moschatellina L., Saussurea dubia Freyn, Petasites frigidus (L.) Fries. Моховой покров часто выражен, развит, покрытие нередко достигает 70-80%. Доминирует Rhytidium rugosum (Hedw.) Kindb., роль Hylocomium splendens (Hedw.) B.S.G. var. splendens снижена. Присутствуют напочвенные и эпифитные лишайники. Часто наблюдается надежное возобновление ели. Сильная редукция кустарникового и травяного покровов объясняется сильными эдификаторными свойствами елового древостоя.
Естественный сукцессионный ряд лесной и кустарниковой растительности долины Средней Лены закономерно заканчивается фазой лиственничных лесов, наступление которой связано, главным образом, с повышением уровня мерзлоты и связанными с этим изменениями гидротермических свойств почвогрунта [1; 4; 5]. Как известно, на водораздельных пространствах и высоких надпойменных террасах Центральной Якутии лиственничные леса являются климаксовыми, зональными формациями. На последних стадиях характерно появление и господство зональной растительности – лиственничников с лишайниково-моховым покровом и травяно-кустарничковым ярусом из виолентных вечнозеленых, длиннокорневищных мезоолиготрофных (Vaccinium vitis-idaea) и олиготрофных лесо-болотных с глубокой корневой системой (Ledum palustre) криофитных кустарничков с длительным парциальным циклом развития. Это диктуется зональным климатом, по мере того как в высоких надпойменных террасах смягчающее влияние реки затухает, начинают господствовать параметры зонального климата, а также достигает своего развития почвенный профиль, стабилизируется мерзлота и достигает средних значений, приближающихся к значениям на водоразделах.
Выводы
Таким образом, в ходе сукцессионной динамики состав и структура растительных сообществ закономерно трансформируется с изменением внешних условий. Сукцессионные смены в долине Средней Лены демонстрируют наличие общих принципиальных признаков, присущих аллювиогенной динамике в долинах крупных рек – векторное качественно-структурное усложнение экосистем в процессе перехода от открытых беспокровных ивовых группировок к высокосомкнутым кустарниковым сообществам и березовым, еловым лесам, затем к зональным лиственничникам. В напочвенном покрове на первых этапах сукцессий появляются пионерные эксплеренты, одно-, двулетники, хвощ полевой, затем господствуют многолетние крупные злаки с утолщенными длинными корневищами и гигромезофильные растения с ползучими столоновидными побегами. На средних стадиях основное ядро видов составляют рыхло- и плотнодерновинные злаки и представители ксеромезофильного, мезоксерофильного лугового, степного разнотравья с участием эутрофного и кальцефильного высокотравья. На последних стадиях характерно появление лишайниково-мохового покрова как структурного элемента и господство в травяно-кустарничковом ярусе виолентных вечнозеленых, длиннокорневищных мезоолиготрофных (Vaccinium vitis-idaea) и олиготрофных лесо-болотных (Ledum palustre) кустарничков с длительным циклом развития. В целом вдоль динамической серии наблюдается тренд ксеромезофитизации и криофитизации, что связано с затуханием пойменного режима, повышением уровня сезонно-талого слоя и нарастающим диктатом зонального резко континентального климата.
Исследования выполнены в рамках проектов НИР ИБПК СО РАН № 52.1.11 «Разнообразие растительного мира таёжной зоны Якутии: структура, динамика, сохранение», № 52.2.8 «Лесные экосистемы криолитозоны Якутии в условиях глобального изменения климата и антропогенного воздействия: состав, структура, продуктивность, прогноз динамики».