



В составе хвойных растений, рекомендуемых для озеленения, хвойные виды р.Picea занимают одно из первых мест, как основные лесообразующие породы, и благодаря своей декоративности, исключительной неприхотливости и долголетию ели на многие годы создают особый колорит и декоративность ландшафта в озеленении.
Обязательным условием успешности фотоассимиляционных процессов хвойных является оптимальное для каждого вида количество ФАР. Низкий уровень освещенности является критическим стрессором, влияющим на ростовые процессы ряда ювенильных древесных растений. Ранее нами установлены ответные реакции саженцев ели при 50 % ФАР [1,4]. У лиственных древесных растений в модельных экспериментах 95 % затенение всходов дуба черешчатого вызывало полную гибель всходов дуба к началу второго периода вегетации [6].
Цель данной работы состояла в оценке показателей водного обмена трех видов ели в условиях низкого уровня ФАР в вегетационных опытах.
Объекты и методы
Вегетационный опыт был заложен в мае 2009 г. в условиях летней теплицы на Горнотаежной станции и продолжался в течение 2010 и 2011 гг. Объекты исследований 3–5-летние саженцы ели аянской (Piceaajanensis Lind. et. Gold.), ели корейской (Piceakorainensis Nakai) – местные виды и ель обыкновенная (Piceaabies (L.) Karst.) – интродуцент. Вегетационные опыты проводили в однотипных почвенных и климатических условиях. Исходная влажность почвы 65–70 %. В опытном варианте использовали притенение части опытных растений под пологом для снижения освещенности в течение от 30 до 60 дней. В контроле рост саженцев под обычной полиэтиленовой пленкой вегетационного домика. Интенсивность транспирации хвои определяли методом быстрого взвешивания [3]. Относительное содержание воды (ОСВ) определяли на изолированной хвое, помещенной в закрытые чашки Петри на дистиллированной воде до полного насыщения тканей хвои в течение 24 часов и рассчитывали по формуле: ОСВ (%) = (В сыр.– В сух.)/(Втур.-В сух.) х 100 [8]. ОПВ определяли по методике [9]. Измерение параметров микроклимата: температуры и влажность воздуха, ФАР и УФ радиации проводили подекадно – 5, 15 и 25 числа каждого месяца на приборе ТКА-ПКМ-42.
Результаты
В 2009 и 2011 гг. температура воздуха в контрольном варианте находилась в пределах 28–33 ºС, а в опыте с притенением несколько меньше, чем в контроле, всего 24–28 ºС. По микроклиматическим параметрам 2010 г. характеризуется ранней весной и аномально жарким летом. Уже в июне месяце 2010 г. температура воздуха достигала 39,1 ºС, а в отдельные дни и более 40 ºС. Влажность воздуха в летние месяцы (июнь-июль) находилась на уровне 33.2–42,6 % в контроле и 35,1, а в опыте с притенением – 48,5 %.
Освещенность (ФАР) под пленкой вегетационного опыта (контроль) в 2009 и 2011 гг. в июне – июле соответствовала 337(х 100) – 577(х 100) лк, а в варианте опыта с притенением всего 104(х100) – 416(х 100)лк (табл.). В 2010 г. освещенность в контроле значительно выше, чем в предыдущий и последующий годы, и соответствует 790 – 893(х100) лк в контроле и 77 – 382(х100) лк в опыте. В опыте с притенением освещенность под пологом по отношению к контролю по средне декадным данным соответствует в летний период 2009 г. 27 %; 2010 г. –17 %; 2011 г. – 8 %.
Показатели УФ-радиации в июне 2009 г. в контрольных условиях соответствовали 190-283(х100) мвт/м2, в притенении – 42-48(х100) мвт/м2, а в июне-июле 2011 г. всего 4(100)-6(100) мвт/м2. В 2010 г. значения УФ-радиации были более высокие и соответствуют 249-312(х100) мвт/м2 в июне-июле в контрольном варианте и 85(х100)-225(100) мвт/м2 в опыте.
В 2009 г. показатели интенсивности транспирации хвои саженцев в опыте и в контроле в летний период вегетации находятся у разных видов в пределах для ели аянской 190–330 мг/г в час, ели корейской 166–405 мг/г, ели обыкновенной 44–330 мг/г в час. В сезонной динамике транспирации хвои ели аянской и ели обыкновенной наблюдали максимум в июле, а у ели корейской он смещен к третьей декаде августа. У всех трех видов ели в течение лета 2009 г. интенсивность транспирации больше в варианте опыт, чем в контроле (рис. 1 а).
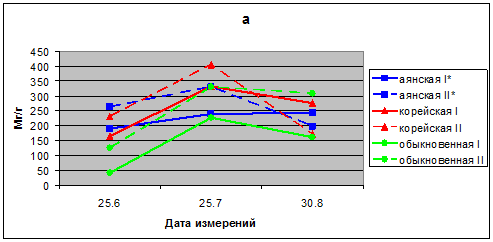
Рис. 1. а. Интенсивность транспирации хвои видов р. Picea в вегетационных опытах (мг/г сырого веса в час); а – 2009; I* – контроль; II* – опыт
В 2010 г. значения транспирации хвои с июля по сентябрь для саженцев ели аянской соответствуют 166–570 мг/г в час, ели корейской 149–346, ели обыкновенной 153–322 мг/г в час. Максимальная величина транспирации – 570 мг/г отмечена в хвое ели аянской в середине августа в опытном варианте (рис. 1б).
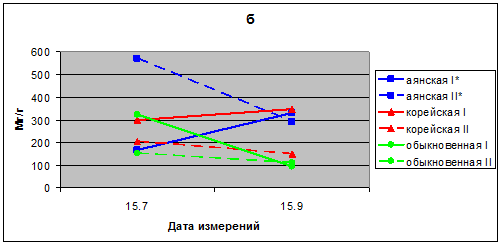
Рис. 1б. Интенсивность транспирации хвои саженцев ели в вегетационном опыте 2010 г.; I* – контроль; II* – опыт
В 2011 г. у исследуемых видов показатели интенсивности транспирации соответствует у ели аянской 141–235 мг/г, ели корейской 115–159 ели обыкновенной 77–160 мг/г в час. В июле этого г. интенсивность транспирации у саженцев в опыте больше, чем в контроле.
Транспирационная способность хвои интродуцента ели обыкновенной ниже, чем у местных видов – ели аянской и корейской, особенно в начале вегетации. В мае месяце в опыте и контроле транспирация хвои всего 44–45 мг/г (рис. 1в).
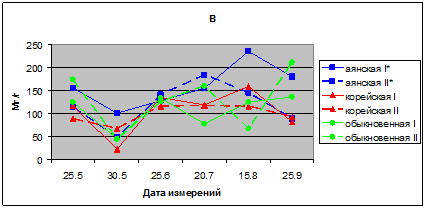
Рис. 1в. Интенсивность транспирации хвои видов р. Picea в вегетационных опытах (мг/г сырого веса в час); в – 2011. I* – контроль; II* – опыт
В 2009 г. относительное содержание воды (ОСВ) в хвое находится в пределах 86,3–97,0 % в хвое ели аянской, 84,6–95,3 % в хвое ели корейской, и 85,6–96,0 у саженцев ели обыкновенной. В летний период вегетации ОСВ только в хвое ели аянской в опыте больше, чем в контроле. ОСВ в хвое двух других видов ели, напротив, у контрольных растений несколько больше, чем у опытных. В сезонной динамике у всех видов ели имеется тенденция снижения уровня ОСВ в хвое к сентябрю месяцу (рис. 2а).
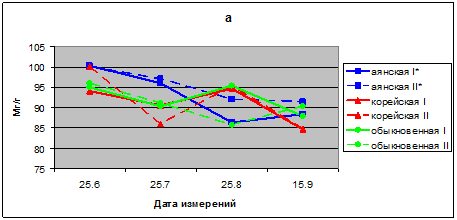
Рис. 2а. Относительное содержание воды в хвое видов р. Picea в условиях низкого уровня освещенности, %. а – 2009; б – 2010; в – 2011, I* – контроль; II* – опыт
В июле и сентябре 2010 г. ОСВ в хвое всех исследуемых видов находится в пределах 83,5–91, 4 %. Показатели ОСВ в хвое ели аянской и обыкновенной больше в опыте, чем в контроле. У саженцев ели корейской ОСВ контроле и в опыте имеют близкие значения. В сезонной динамике наблюдается небольшое увеличение ОСВ к сентябрю месяцу и в опыте и в контроле (рис. 2 б).
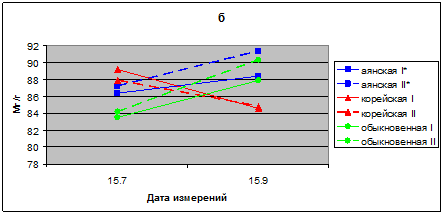
Рис. 2 б. Относительное содержание воды в % в хвое видов р. Picea
в условиях низкого уровня освещенности в 2010 г. I* – контроль; II* – опыт
В 2011 г. у всех видов ели ОСВ меньше, чем в предыдущие годы, и соответствуют 71,0–91,6 % в хвое ели аянской, 67,0–85,6% ели корейской и 61,9 83,6 в хвое ели обыкновенной. В основном ОСВ хвои всех видов ели находится в пределах от 60 до 85 %, с небольшой разницей значений ОСВ между опытом и контролем (рис. 2в).
Показатели относительной потери воды (ОПВ) в летний период 2009 г. соответствуют 0, 40–0,52 мг/г, и только в хвое ели обыкновенной в контрольном варианте значения ОПВ были самыми низкими 0,15 мг/г в час (рис. 3а).
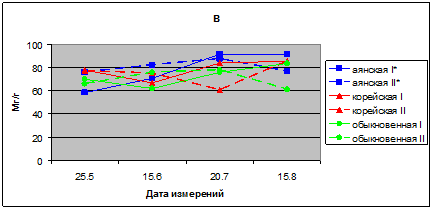
Рис. 2 в. Относительное содержание воды, (%) в хвое видов р. Picea в 2011 г.
I* – контроль; II* – опыт
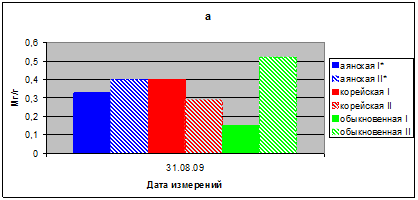
Рис. 3 а. ОПВ (мг/г сыр.веса в час) в августе 2009 г.
Показатели ОПВ хвои в летний период 2010 г. значительно выше, чем в 2009 г., и соответствуют 0,76–2,54 мг/г. Скорость потери воды у саженцев в опыте больше, чем в контроле. Максимальные значения ОПВ отмечены в августе месяце, когда значения ОПВ у саженцев ели аянской в контроле достигают 2.52 мг/г, а в опыте – 3,45 мг/г в час. ОПВ в хвое видов ели корейской и ели обыкновенной имеют близкие значения (рис. 3 б).
Показатели ОПВ в 2011 г. у саженцев всех изучаемых видов ели находятся в пределах 0,031–0,19 мг/г. Максимальные показатели ОПВ хвои у всех видов ели в мае месяце соответствуют 0,11–0,19мг/г в час. Со второй декады июня наблюдается снижение относительной потери воды в хвое всех саженцев ели (рис. 3 в).
В сезонной динамике ОПВ хвои саженцев ели в 2011 г. прослеживается тенденция снижения показателей ОПВ с 2009 г. к сентябрю месяцу 2011 г. (рис. 3в). Показатели влажности почвы в контроле и опыте варьировали от 15 до 45 %. По сравнению с 2009 и 2011 годами наблюдается значительное снижение влажности почвы в 2010 г., когда минимальные значения влажности почвы были в пределах 13 %. Значения влажности почвы, как правило, больше в опыте, чем в контроле.
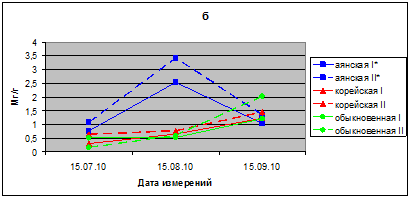
Рис. 3б. ОПВ (мг/г сырого веса в час) в хвое саженцев ели в летний период вегетации 2010 г.
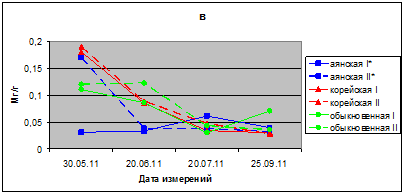
Рис. 3 в. ОПВ (мг/г сырого веса в час) в хвое видов р. Picea в вегетационных опытах 2011 г.
I* – контроль; II* – опыт
Все виды саженцев ели различаются по оводненности хвои в пределах от 47,5 до 73,7 %. Значения влажности хвои имеют тенденцию снижения ее показателей с 2009–2010 г. к 2011 г. Влажность хвои в 2011 г. в 2,4–2,9 раза меньше, чем в 2009 г. и в 2,4–3,5 раза меньше, чем в 2010 г. В контроле и в опыте показатели влажности хвои видов ели располагаются в убывающем порядке: ель обыкновенная, ель корейская, ель аянская (рис. 4).
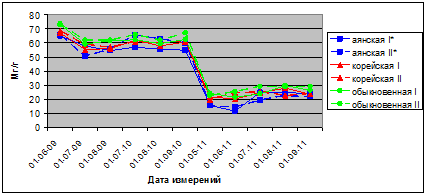
Рис. 4. Влажность хвои саженцев ели в условиях притенения в вегетационных опытах
2009–2011 гг.
Обсуждение результатов
На основании полученных данных установлено, что транспирационная способность у всех видов ели значительно ниже в начале вегетации, по сравнению с летним периодом. Интенсивность транспирации хвои у местных видов – ели аянской и ели корейской, в мае и июне месяцах соответствует 115–190 мг/г в час, а в июле и августе – 330-450 мг/г в час, достигая у саженцев ели аянской максимального уровня 570 мг/г. В отличие от местных видов, интродуцентель обыкновенная в начале вегетации имела очень низкие значения транспирации хвои – всего 41–77 мг/г в час, что обусловлено особенностью ритмов развития этого вида ели в условиях юга Приморья. Все виды ели имели более низкую интенсивность транспирации в 2011 г., по сравнению с 2009 и с 2010 г. Интенсивность транспирации хвои саженцев всех видов в опытном варианте с притенением превышает ее значения в контрольных условиях. Транспирация хвои в 2011 г. в 1,5–3,0 раза меньше, чем в 2010 г. У взрослых хвойных деревьев в лесном массиве дендрария ГТС, в летний период вегетации в августе месяце интенсивность транспирации соответствует: у ели корейской 150, ели аянской – 240 мг/ час [7]. ОСВ хвои соответствует по средним данным в 2009 г. – 96 %, в 2010 – 87 %, а в 2011 г. – 77 %. В 2011 г. ОСВ хвои ели аянской на 14,4 %, а ели обыкновенной на 22,2 % в опыте меньше, чем в контроле. ОПВ в 1,4–1,7 раза в условиях опыта больше, чем в контроле.
Наиболее точно водный статус растений характеризует относительное содержание воды (ОСВ), отражающее степень насыщенности тканей водой по сравнению с их полной тургесцентностью. Уровень относительного содержания воды ОСВ в хвое является показателем стабильности водного обмена растений. В 2011 г. ОСВ хвои саженцев у всех видов ели значительно меньше, чем в предыдущие годы. По данным в 2009 г. 96 %, в 2010 г. – 87 %, а в 2011 г. – 77 %. В 2011 г. ОСВ хвои ели аянской на 14,4 % , а ели обыкновенной на 22,2 % меньше в хвое саженцев в опытном варианте, чем в контроле.
2009 г. величина относительной потери воды (ОПВ) в хвое местных видов ели имела близкие значения, 0,30–0,40 мг/г в час. ОПВ хвои ели обыкновенной в среднем соответствует 0,52 мг/г. По сравнению с 2009 и 2011 г. летний период 2010 г. характеризуется более высокими значениями ОПВ, до 2,54 мг/ г в контроле и 3,45 мг/г в час в опыте, что в значительной мере обусловлено климатическими особенностями этого года. Величина ОПВ только в начале вегетации в мае 2011 г. соответствовала 0,11–0,19 мг/г. С июня по сентябрь ОПВ хвои саженцев всех видов на порядок меньше, чем в летние месяцы предыдущих лет. В опытном варианте с притенением скорость потери воды из хвои больше, чем в контрольном варианте, особенно у саженцев интродуцента ели обыкновенной. В сезонной динамике ОПВ имеется тенденция снижения величины ОПВ с 2009 г. к 2011 г., со значительным максимумом скорости потери воды летом аномального по погодным условиям 2010 г. В варианте вегетационных опытов под влиянием значительного (стрессового) снижения ФАР для всех видов саженцев ели характерно снижение показателей водного обмена: интенсивности транспирации, ОСВ и ОПВ хвои. При этом в вариантах опытов с притенением наблюдается и снижение количества пигментов по их сумме в хвое всех изучаемых видов ели в 1,1–1,4 раза.
Выводы
Влияние низкого уровня ФАР в опыте сопровождается снижением транспирационной способности хвои изучаемых видов ели. Интенсивность транспирации у всех видов ели в опыте была меньше, чем в контрольном варианте. Скорость транспирации хвои ели имеет низкие значения в начале вегетации и увеличивается к августу месяцу. Интенсивность транспирации хвои интродуцента ели обыкновенной значительно ниже, чем у местных видов ели.
Анализ сезонной динамики ОСВ показал тенденцию снижения этого показателя с 2009 г. к 2011 г. с 96 % до 77 %. ОСВ тканей хвои ели на 15,4 % меньше в 2011 г., чем в предыдущие годы. Реальный водный дефицит, который испытывали саженцы в этом году, был очень сильным. Падение ОСВ составило от 14,4 до 22,2 % в августе 2011 г. в опытном варианте. Из всех видов ели наименьшие значения ОСВ были в хвое ели обыкновенной: всего 83,6 % в контроле и 61,4 % в опыте.
Относительное содержание воды (ОСВ) у саженцев всех видов ели имеет минимальные значения в начале вегетации (май) и увеличивается к середине лета (август). В годовой динамике наблюдается снижение скорости потери воды (ОПВ) хвои у всех видов ели с 2009 к 2011 г., с максимумом в августе аномально жаркого лета 2010 г. и только у саженцев ели аянской достигает значений 2,54 в контроле и 3,42 мг/г в час в опыте. У саженцев под пологом (опыт) значение ОПВ больше, чем в контроле.
Интродуцент – ель обыкновенная – имеет видоспецифичность формирования водного статуса и ритма развития в условиях Приморья в начале вегетации, но достигает по ростовым индексам и водному обмену аборигенных видов уже в июле, что позволяет рассматривать этот вид ели для введения в культуру лесопосадок и озеленения.
Моделированный нами, постепенно снижающийся к третьему году исследований уровень ФАР (8–10 % от контроля), вызывает временное нарушение показателей водного статуса, восстанавливающихся в репарационный период, что указывает на высокую адаптивную пластичность изучаемых видов ели.