



В последние десятилетия эпифизарный гормон мелатонин подвергался многочисленным и всесторонним исследованиям. Экспериментально установлены синхронизирующие свойства мелатонина в организации циркадианного ритма двигательной активности [5], ритма сна-бодрствования [1], показаны его свойства как протектора ДНК и сильного антиоксиданта [4, 6]. Тот факт, что мелатонин является необходимым звеном регуляции циркадианных ритмов различных функций организма, позволяет предполагать его участие и в регуляции функционирования мужских гонад. Последнее должно осуществляться непосредственным или опосредованным влиянием мелатонина на процессы репродукции, дифференцировки и функционирования сперматогенных и эндокринных клеток семенников. Однако вопрос о роли мелатонина в регуляции активности мужских гонад остается открытым. В предыдущих работах мы показали патоморфологические изменения, возникающие в гонадах после эпифизэктомии [3]. Целью настоящей работы явилось исследование компенсаторных возможностей паренхимы семенников эпифизэктомированных белых крыс на фоне введения мелатонина.
Материалы и методы. Опыты выполнены на 270 самцах беспородных белых крыс массой 160-200г. Животные в течение 20 дней адаптировались к режиму освещенность: темнота=12:12 (освещение с 6 до 18 ч). На всем протяжении опыта доступ к пище и воде был свободным. По истечении адаптационного периода крысы были разделены на три экспериментальные группы: интактные контрольные (n=90), эпифизэктомированные (n=90), эпифизэктомированные с последующим введением мелатонина (n=90). Все эксперименты, уход и содержание животных осуществлялось в соответствии с Дерективой №63 от 22.09.10г. Президиума и Парламента Европы «О защите животных, используемых для научных исследований». Животным 2 и 3 группы была проведена операция эпифизктомии по оригинальной методике [2].
Животным 3 группы ежедневно с 26-го по 41-й день после эпифизэктомии в 18 часов (время смены фоторежима со световой фазы на темновую) подкожно вводили свежеприготовленный раствор мелатонина (в дозе 10мг/кг), что моделировало ночной подъем уровня мелатонина в крови. Забор гистологического материала производили на 40 – 41-й день после эпифизэктомии через каждые три часа в течение двух суток, что обеспечивало исследование функциональной активности семенников на протяжении двух периодов циркадианного ритма.
Семенники фиксировали в жидкости Карнуа и готовили парафиновые срезы 5-6мкм по стандартной гистологической методике. Срезы окрашивали реакцией с Шифф-йодной кислотой (ШИК-реакция), ядра клеток доокрашивали гематоксилином Майера. У животных каждой группы определяли абсолютную массу семенников (г), суточную динамику митотического индекса сперматогоний промежуточного типа и типа Б (‰), частоту встречаемости метафазы мейоза и спермиации (‰), количество активных клеток Лейдига на 1000 клеток интерстиция, общую функциональную активность перитубулярных эндокриноцитов (суммарная площадь сечения ядер всех выявленных активных эндокриноцитов из 1000 клеток интерстиция) в каждой временной точке.
Для всех параметров при статистической обработке вариационных рядов вычислялись значения средней арифметической взвешенной (М), ошибки средней арифметической взвешенной (m). Достоверность различий между показателями оценивалась t-критерием Фишера-Стьюдента. Уровень значимости различий был принят р<0,05. Для оценки наличия ритма применяли регрессионный анализ методом наименьших квадратов и спектрального анализа.
Результаты исследования и их обсуждение. После введения мелатонина эпифизэктомированным самцам белых крыс однократно в течение суток на протяжении двух недель общая организация семенников морфологически не отличалась от интактных и эпифизэктомированных животных. Во всех извитых семенных канальцах сохранялся активный сперматогенез (рис. 1).
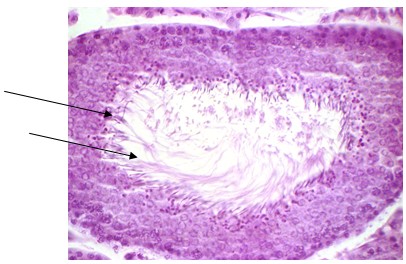
Рис. 1. Извитые семенные канальцы на стадии спермиации у эпифизэктомированных белых крыс после введения мелатонина. Окрашивание: ШИК-реакция, с доокраской ядер гематоксилином, увеличение х 400. Наблюдается выход сперматозоидов в просвет извитого семенного канальца
Площадь, занимаемая интерстицием, составила 9,84±3,14% от общей площади поперечного сечения семенника. По сравнению с интактными и эпифизэктомированными животными достоверного изменения соотношения сперматогенная/эндокринная ткань не наблюдалось, что свидетельствует о сохранении среднего уровня активности течения сперматогенеза. Количественное отношение и взаимное расположение сперматогенных клеток не отличалось от группы интактных контрольных животных, в каждом извитом канальце определялись цитологический профиль и стадия сперматогенного цикла.
Однократные в течение суток инъекции мелатонина подавляли высокую митотическую активность сперматогоний, наблюдавшуюся после эпифизэктомии только на короткое время, а затем митотический индекс достигал прежних, высоких значений. Среднесуточный уровень митотической активности сперматогоний промежуточного типа превышал значения интактных животных на 9,2%, сперматогоний типа Б – на 18,8 %. При графически-параметрическом анализе суточной динамики митотического индекса сперматогоний изученных типов в данной экспериментальной группе отмечалось отсутствие циркадианных ритмов (рис. 2).
Сглаженная кривая динамики митотической активности сперматогоний промежуточного типа характеризовалась кривой без ритмических колебаний и не обнаруживала связи со свето-темновым циклом (рис. 2). Активные фазы наблюдались как в светлое, так и в темное время суток. Максимальные значения показателей фиксировались в 24ч, 9ч и 15 ч первых суток, 24ч, 6ч и 12ч вторых суток эксперимента. Абсолютная амплитуда составила 38,32‰, что практически не отличалось от показателей у интактных животных. Средние значения митотического индекса в ночные и дневные часы достоверно не различались.
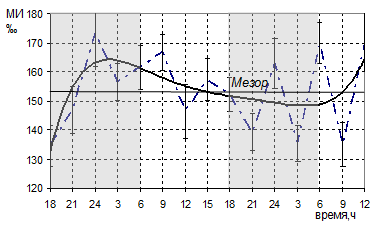
Рис.2. Суточная динамика митотической активности сперматогоний промежуточного типа эпифизэктомированных белых крыс после введения мелатонина
Обработка полученных данных методом спектрального анализа показала наличие колебаний с периодом 24 ч для сперматогоний промежуточного типа, однако амплитуда колебаний была незначительна. Для сперматогоний типа Б выявлены колебания с периодом 17 ч, что характерно для ультрадианного ритма. Анализ митотической активности сперматогоний методом наименьших квадратов выявил наличие ультрадианного ритма митотической активности с периодом 19 ч и инфрадианного ритма с периодом 33,5 ч для сперматогоний промежуточного типа. То есть, циркадианный ритм, выявленный методом спектрального анализа, не подтвердился другими методами математического и графического анализа.
При определении суточной динамики частоты встречаемости этапов мейотического деления сперматоцитов и спермиации в группе эпифизэктомированных крыс с введением мелатонина также отмечалось исчезновение циркадианных ритмов их встречаемости.
Таким образом, ежесуточное введение мелатонина эпифизэктомированным животным в течение двух недель не привело к формированию суточных ритмов течения сперматогенеза. Показатели течения сперматогенеза, изученные в данной группе, практически не отличались от показателей в группе эпифизэктомированных животных.
Введение мелатонина эпифизэктомированным животным не повлияло на среднесуточное количество активных клеток Лейдига (760,47±4,89). Суточная динамика, как и в группе эпифизэктомированных животных, характеризовалась кривой без определенной взаимосвязи со свето-темновым циклом (рис.3). Абсолютная амплитуда колебаний составила 58,16 единиц, что превышало значения у интактных и эпифизэктомированных животных. Средние показатели в темное и светлое время суток не имели достоверных различий (752,92±9,19 и 763,17±5,73 соответственно, р>0,05). Метод спектрального анализа не выявил периодических колебаний количества активных гландулоцитов в течение суток. Ультрадианный ритм с периодом 6,5 ч выявлен только методом наименьших квадратов.
Суточная динамика суммарной площади сечения ядер активных клеток Лейдига в группе эпифизэктомированных животных, получающих ежесуточно мелатонин, также не имела циркадианной ритмичности. Профили синусоиды, на графике суточной динамики не имели связи со свето-темновым циклом, достоверное повышение активности наблюдалось только во вторые сутки эксперимента, причем как в темное, так и в светлое время суток (24ч и 12ч соответственно) (рис.3). Амплитуда колебаний составила 4096,83 единиц (22,65%). Средние значения в светлое время не отличались от значений, полученных для темного времени суток (15476,94±640,38 мкм2 и 15819,67±474,77мкм2 , р>0,05).
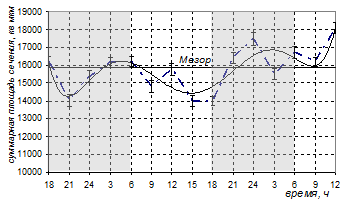
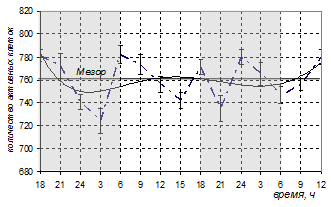
Рис. 3. Суточная динамика количества активных клеток Лейдига и суммарной площади сечения их ядер у эпифизэктомированных белых крыс после введения мелатонина
Математические методы анализа временной динамики активности гландулоцитов семенника также не выявили циркадианной ритмичности изучаемых показателей. Так, метод спектрального анализа показал отсутствие каких-либо ритмических колебаний. Методом наименьших квадратов определены только ультрадианные колебания с периодом 10,3 ч.
В настоящее время мелатонин рассматривается как основной гормон эпифиза. Эффекты мелатонина показывают существенную зависимость от времени суток и сезона года, когда вводился препарат, а также от дозы, режима и способа введения. Регуляторное воздействие мелатонина на репродуктивную систему выявлено для некоторых моноэстричных животных, причем определяющими факторами в данном случае, помимо дозы и длительности введения, являлась фаза репродуктивного цикла, в которую вводился мелатонин. Так, введение мелатонина в вечернее время пальмовой белке вызывало угнетение активности семенников [7]. Показано, что медленное введение мелатонина (подкожный имплант) в течение 47-71 дней не изменяет качества спермы баранов, но увеличивает выработку тестостерона [8].
В проведенном нами исследовании мелатонин вводился при смене световой фазы фотопериода на темновую однократно в течение суток, что должно было смоделировать ночной подъем уровня мелатонина у интактных животных. Однако введение мелатонина эпифизэктомированным животным в нашем эксперименте не привело к восстановлению циркадианного ритма активности мужских гонад. Нарушения функции сохранялись как со стороны герминативной ткани семенников, так и со стороны эндокринной. Используемая нами схема введения мелатонина не обеспечила его длительный антипролиферативный эффект на сперматогонии. Имеются сведения, что после введения экзогенного мелатонина через рот или внутривенно он быстро метаболизируется в печени. Вероятно, однократное в течение суток введение мелатонина не позволило установить достаточный уровень концентрации гормона в крови вследствие его быстрого выведения, что и объясняет отсутствие эффекта мелатонина на формирование циркадианного ритма активности мужских гонад.
Заключение. Таким образом, однократное в течение суток введение мелатонина эпифизэктомированным животным на протяжении двух недель не позволило сформировать циркадианные ритмы течения сперматогенеза и активности эндокринной ткани семенников. Выявленные в этой группе параметры соответствовали показателям, полученным в группе эпифизэктомированных животных. Отмеченное выше отсутствие эффекта однократного в течении суток введения мелатонина на формирование циркадианного ритма пролиферации может рассматриваться как обоснование необходимости разработки форм и способов обеспечения пролонгированного действия мелатонина.
Рецензенты:
Говорухина А.А., д.б.н., зав. кафедрой медико-биологических дисциплин и безопасности жизнедеятельности, ведущий научный сотрудник Лаборатории «Здоровый образ жизни и охрана здоровья» Сургутского государственного педагогического университета, г. Сургут.
Смолькина А.В., д.м.н., доцент, профессор кафедры госпитальной хирургии, анестезиологии, реаниматологии, урологии, травматологии, ортопедии медицинского факультета им. Т.З. Биктимирова Института медицины, экологии и физической культуры ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск.