



Поперечное деление центромеры (misdivision) в мейозе проявляется обычно при гемизиготном состоянии хромосомы и является характерным для моносомных линий. При эуплоидном состоянии генотипа подобное явление обычно не наблюдается. Однако его проявление у анеуплоидов требует своего объяснения, так как оно может быть связано с механизмом реализации общей мейотчсеской программы. Касаясь данной стороны вопроса, следует отметить, что в норме остаются незамеченными для наблюдателя очень многие мейотические процессы. Для их изучения уникальным объектом, кроме мутантных форм, являются серии моносомных линий, созданные по аллополиплодным видам растений [6–8]. Тем не менее предоставленная возможность так и не была до конца использована.
Материал и методы исследования
Исследования проводили на полной серии моносомных линий сорта яровой мягкой пшеницы Мильтурум 553, созданной путем беккроссирования (8-9 беккроссов) исходной моносомной серии сорта Чайниз Спринг (авторы: Цильке Р. A. , Цильке И. А. , Жарков Н. А., Присяжная Л. П. ). Потомство моносомных растений высевали в теплице, в сосуды и выращивали при 16 часовом освещении с 23/18о С температурным циклом.
Фиксацию микроспороцитов осуществляли в межфазный период выход в трубку – колошение смесью Ньюкомера. Для этого с каждого растения брали по одному колосу. Цитологический анализ мейоза проводили на временных давленых ацетокарминовых препаратах с помощь светового микроскопа МБИ-3 при увеличении 20х10. По каждой линии анализировали от 10 до 15 растений. Для цитологического анализа использовались только первый и второй цветок колосков, расположенных в нижней части колоса. По результатам исследований был проведен корреляционный анализ [2].
Результаты исследования и их обсуждение
По результатам проведенных исследований в анафазе I мейоза наблюдалось три основных типа поведения унивалента. Первый и преобладающий тип характеризуется униполярной ориентацией унивалента, при котором моносома отходила к одному из противоположных полюсов синхронно с остальными хромосомами, входящими в метафазе I в состав бивалентов. Данный тип поведения унивалента может трактоваться как мейотический, или как униполярный. В этом случае, при нормальном синапсисе гомологичных пар, анафазные клетки не имели отстающих элементов.
Второй тип связан с биполярной ориентацией центромеры, приводящей к продольному расщеплению унивалента. В связи с тем, что деление хромосомы в данном случае проходит по типу митоза, то такое поведение унивалента было интерпретировано как митотическое, или как биполярное.
Третий тип поведения унивалента может быть охарактеризован как аполярный, поскольку очень часто наблюдались случаи, когда моносома не взаимодействовала с полюсами. Сохраняя свою целостность до конца анафазы, она давала начало формированию микроядра в диадах. Коэффициент парной корреляции между числом анафазных клеток с целой отстающей хромосомой и количеством диад с микроядром составил 0,71. При расчете множественной корреляции, где кроме двух параметров учитывалось наличие анафазных клеток с одной отстающей хроматидой, частный коэффициент был близок к единице (r = 0,93).
Кроме продольного расщепления унивалента в анафазе первой относительно часто наблюдалось поперечное деление центромеры (misdivision) одной или сразу двух составляющих его хроматид (рис.1).
а)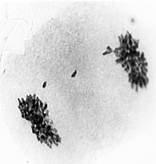 б)
б)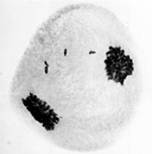 в)
в)
Рис. 1. Поперечное деление центромер унивалентной хромосомы в первой анафазе мейоза: а – одной хроматиды; б – двух хроматид; в – без продольного расщепления унивалента. Ув. 60 х 10
По классификации, разработанной Сирсом [9], у анализируемого материала встречались все выделенные им классы, кроме тех случаев, когда четыре плеча ориентированы к одному полюсу (класс «d»). У серии моносомных линий Мильтурум 553 чаще всего наблюдалось поперечное деление центромеры одной из сестринских хроматид без продольного расщепления унивалента (рис.1, в). В целом, по серии моносомных линий их количество составило 0,65 %. Данный тип поперечного деления центромеры унивалента может быть выделен в отдельный класс «е». По частоте встречаемости второе место занимают случаи поперечного деления одной хроматиды с правильным расхождением образовавшихся телоцентриков (плеч хроматиды). Реже встречались конфигурации классов «в» и «с». Частота поперечного деления унивалента существенно варьировала по линиям (таблица 1).
Таблица 1. Частота (%) биполярной ориентации унивалента и поперечного деления его центромеры в анафазе I мейоза у серии моносомных линий сорта яровой мягкой пшеницы Мильтурум 553
|
Хромосома |
Изучено клеток |
Количество клеток, % |
|
|
с биполярной ориентацией унивалента |
с поперечным делением центромеры унивалентной хромосомы |
||
|
1A |
1274 |
27,48 |
3,06 |
|
2A |
2081 |
10,14 |
0,48 |
|
3A |
1953 |
33,90 |
1,33 |
|
4A |
1751 |
34,27 |
3,31 |
|
5A |
1572 |
36,64 |
3,37 |
|
6A |
1756 |
27,11 |
2,73 |
|
7A |
1432 |
47,55 |
0,28 |
|
1B |
1069 |
30,78 |
1,96 |
|
2B |
1243 |
46,82 |
3,22 |
|
3B |
1444 |
29,99 |
1,32 |
|
4B |
1176 |
32,34 |
1,02 |
|
5B |
1337 |
35,00 |
1,35 |
|
6B |
1193 |
17,68 |
2,35 |
|
7B |
1090 |
27,16 |
2,75 |
|
1D |
1084 |
12,02 |
0,55 |
|
2D |
1624 |
15,15 |
0,25 |
|
3D |
1148 |
26,92 |
0,60 |
|
4D |
1434 |
30,62 |
1,12 |
|
5D |
1348 |
39,68 |
2,60 |
|
6D |
1540 |
30,51 |
1,17 |
|
7D |
1610 |
23,17 |
0,50 |
|
Геном А |
11819 |
30,22 |
2,08 |
|
Геном В |
8552 |
31,64 |
2,00 |
|
Геном D |
9788 |
25,47 |
0,97 |
|
X средняя |
30159 |
29,10 |
1,68 |
По данным цитологического анализа чаще процессу поперечного деления подвергались центромеры унивалента по хромосомам 5А (3,37 %), 4А (3,31 %), 2В (3,22 %) и 1А (3,06 %). Реже – 2D (0,25 %), 7A (0,28 %) и 2А (0,48 %). В целом хромосомы генома D в гемизиготном состоянии реже подвергались поперечному делению, чем геномов А и В (таблица 1). Между частотами биполярной ориентации унивалента и поперечным делением его центромеры проявилась слабая положительная корреляционная зависимость (r = 0,38).
По результатам проведенных исследований акт поперечного разрыва центромеры унивалента у серии моносомных линий Мильтурум 553 наблюдался значительно реже, чем это отмечалось Сирсом [9] в отношении аналогичной серии сорта Чайниз Спринг. Возможно, более низкий уровень misdivision в микроспороцитах анализируемого материала связан с тем, что часто унивалентная хромосома не взаимодействовала с полюсами, проявляя аполярный тип своего поведения.
В анафазе II большинство клеток не имели отстающих элементов. На их долю приходилось в среднем по моносомной серии около 69 % всех изученных клеток. Редко (1,06 %) отмечались случаи наличия отстающей целой унивалентной хромосомы. Чаще всего в межполюсной зоне наблюдалось присутствие одной или двух хроматид, последние из которых могли быть расположены как в одной, так разных дочерних клетках диады. В некоторых клетках одиночная хроматида, расположенная в экваториальной плоскости, не имела проявления какой-либо заметной полярной ориентации.
В анафазе второго мейотического деления также отмечались случаи поперечного деления центромеры одной или сразу двух хроматид (рис. 2).
а)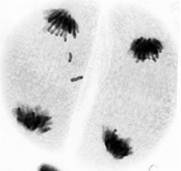 б)
б)
Рис. 2. Поперечное деление центромеры хроматид унивалентной хромосомы в анафазе II мейоза: а – одной хроматиды; б – двух хроматид. Ув. 60 х 10
По результатам проведенного цитологического анализа усредненные показатели частоты поперечного деления центромеры унивалентной хромосомы в анафазе I и анафазе II оказались близки между собой (1,68 и 1,99 % соответственно). Однако характер распределения этих частот по линиям в анафазе второго деления существенно отличался от такового в анафазе I (таблица 2).
Как видно из таблицы 2, чаще поперечному делению подвергались центромеры хроматид унивалентов по хромосомам 1В (6,20 %), 6B (5,01 %), 5D (5,31 %), 7B (3,75 %) и 5A (3,31 %). Реже – 2D (0,31 %), 3D (0,48 %) и 2A(0,60 %). В целом misdivision превалировал в анафазе II при гемизиготном состоянии хромосом генома В (таблица 2). Частота поперечного деления хроматид в анафазе II положительно коррелировала с количеством анафазных клеток с отстающими хроматидами (r = 0,52), что оказалось достоверным при Р ≤ 0,05. Слабая положительная корреляция проявилась между частотой misdivision во второй анафазе и долей клеток с биполярной ориентацией унивалента в анафазе первой (r = 0,25).
В работе J.M Vega.и M. Feldman [10] поперечное деление центромеры унивалента связывают с функцией кинетохора, при которой имеющиеся отдельные его домены способны менять свою связь с полюсами. Однако несколько иное объяснение имеет механизм поперечного деления моносомы, если исходить из ранее предложенной концепции «разводящих нитей», которая была разработана на основе результатов анализа характера поведения унивалента в анафазе I мейоза у серии моносомных линий пшеницы Мильтурум 553 [3]. Суть ее содержания сводится к следующему:
1. Переход хромосом от эквационного деления к редукционному и обратно осуществляется сегрегацией зоны исходного полюса. В качестве последней могут быть: определенный участок оболочки ядра, митотический центр или иное структурное образование.
2. При сегрегации зоны исходного полюса происходит смена полюсной ориентации точек фиксации связующих нитей центромер с полюсом одного гаплоидного набора хромосом путем их перемещения на одну сторону от линии его потенциального раздела. В это время их гомологи сохраняют свою прежнюю биполярную (митотическую) ориентацию. Связь центромер с полюсом также может осуществляться с помощью микрофибрил, микротрубочек или непосредственного контакта. Изменение ориентации хромосом одного гаплоидного набора приводит в соответствие взаимное расположение гомологов и служит пусковым механизмом начала их сближения.
Таблица 2. Частота поперечного деления центромеры хроматид в анафазе II, %
|
Хромосома |
Изучено клеток |
Количество клеток, % |
|
|
С отстающими хроматидами |
С поперечным делением центромеры |
||
|
1А |
1168 |
29,46 |
2,05 |
|
2A |
1496 |
17,05 |
0,60 |
|
3A |
1319 |
26,08 |
0,83 |
|
4A |
1693 |
35,79 |
2,30 |
|
5A |
665 |
35,79 |
3,31 |
|
6A |
1541 |
23,30 |
1,10 |
|
7A |
1335 |
39,92 |
1,05 |
|
1B |
1016 |
32,58 |
6,20 |
|
2B |
935 |
39,04 |
2,78 |
|
3B |
1417 |
28,65 |
1,48 |
|
4B |
1126 |
28,24 |
1,95 |
|
5B |
1267 |
36,31 |
2,21 |
|
6B |
1038 |
28,90 |
5,01 |
|
7B |
1119 |
29,31 |
3,75 |
|
1D |
1258 |
17,96 |
0,95 |
|
2D |
1307 |
15,23 |
0,31 |
|
3D |
1253 |
23,46 |
0,48 |
|
4D |
1334 |
32,61 |
0,67 |
|
5D |
1168 |
47,78 |
5,31 |
|
6D |
1178 |
42,36 |
1,61 |
|
7D |
1368 |
19,23 |
0,88 |
|
Геном А |
9217 |
29,07 |
1,45 |
|
B |
7918 |
31,67 |
3,13 |
|
D |
8866 |
27,90 |
1,42 |
|
X ср. |
26001 |
29,47 |
1,99 |
3. После контакта гомологов и начала их синапсиса хромосомы, которые до сих пор сохраняли свою биполярную ориентацию, утрачивают свою связь с зоной исходного полюса. Именно данным обстоятельством может объясняться наблюдаемое в зиготене состояние «букета». Таким ходом событий достигается сразу две цели. 1. Облегчается прохождение конъюгации хромосом. 2. Сохраняется упорядоченность пространственного расположения хромосом относительно друг друга во время их синапсиса.
4. При завершении конъюгации хромосом связь их центромер с полюсом восстанавливается, но не в прежнем митотическом порядке, а в последовательности, обеспечивающей редукционный тип деления хромосом при разведении кинетохорных нитей веретена и встраивание их в центральное веретено деления.
Таким образом, из вышесказанного следует, что смена полюсной ориентации в переходный период одного гаплоидного набора осуществляется сегрегацией зоны исходного полюса и ему для этого не требуется наличие партнера. Однако второму гаплоидному набора хромосом для ко-ориентации центромер необходимо наличие синапсиса. Согласно полученным экспериментальным данным [3] вероятность того, какой из двух гомологов окажется в том или ином положении, составляет 50:50 %. Именно поэтому унивалент, оказавшись в первом положении, ведет себя по типу редукционного деления, а во втором – эквационного с частой реализацией потенциальной способности утрачивать свою связь с полюсом.
Правомерность ранее выдвинутой концепции «разводящих нитей» подтверждается данными литературы [5] и результатами проведенных собственных исследований, согласно которым в прометафазе мейоза пшеницы в норме наблюдается симметричное, трехполюсное веретено (рис. 3).
а)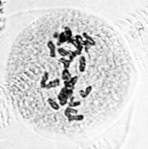 б)
б)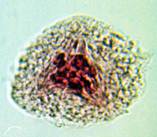 в)
в)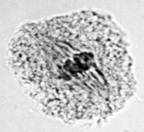
Рис. 3. Процесс формирования веретена при первом мейотическом делении клеток у мягкой пшеницы: а – профаза; б – прометафаза; в – метафаза I. Ув. 60х10
Из представленной на рисунке 3, б конфигурации нетрудно догадаться, что два полюса принадлежат центральному веретену, а третий – кинетохорному. Естественно, исход дальнейших событий будет определяться характером расщепления кинетохорного полюса с последующим разведением и вхождением его нитей в центральное веретено деления.
Поперечное деление центромеры унивалента возникает по причине недостаточно корректного прохождения сегрегации зоны исходного полюса. При полюсной ко-ориентации хромосом одного гаплоидного набора в зону потенциального раздела попадают точки фиксации соединяющих нитей хроматидных центромер той части хромосом, которые в это время сохраняют свое прежнюю биполярную ориентацию. В обычном эуплоидном состоянии подобного рода погрешности достаточно легко устраняются путем прерывания их связи с полюсом и ее восстановления по завершении конъюгации. Совсем другое дело, когда хромосома находится в гемизиготном состоянии и не имеет возможности изменить свое положение.
По описанию, данному К. Босток и Э. Самнера [1], центромера на электронной фотографии имеет в своей зоне две пары плотных телец, разделяющих ее на две хроматиды и четыре плеча. Очевидно, данные участки являются районами потенциального раздела центромеры. Поэтом при подготовке клетки к делению в зоне центромеры формируется кинетохор, который усиливает ее жесткость. При этом фиксация системы микротрубочек должна быть распределена по центромере таким образом, чтобы обеспечить ее целостность при расхождении хромосом в анафазе I и хроматид в анафазе II мейоза. Однако исход событий во многом зависит от полюсной детерминации характера разведения кинетохорных нитей веретена деления. Варианты возможных событий представлены на рисунке 4.
При смене полюсной ориентации одного гаплоидного ряда хромосом довольно часто происходит смещение точек фиксации связующих нитей центромер унивалента, который в это время оказывается в пассивном положении и сохраняет свою прежнюю биполярную ориентацию. Если полюсное расположение фиксации пучка микротрубочек одной из хроматид относительно линии раздела совпадает с линией потенциального деления ее центромеры на два плеча (рис. 4, в), то при его расщеплении в анафазе I будет наблюдаться ситуация, когда унивалент имеет поперечное деление данной хроматиды без его продольного расщепления (класс «е»). В том случае, когда в таком положении окажутся обе хроматиды (рис. 4, г), то в анафазе I должно образоваться две изохромосомы (поперечное деление центромеры унивалента без его продольного расщепления). Проведенные цитологические наблюдения показывают, что расположение пучка микротрубочек в зоне деления может иметь различные варианты. При несовпадении направления расщепления пучка микротрубочек с линией потенциального поперечного раздела центромеры довольно часто возникают ситуации, когда один из образовавшихся телоцентриков в анафазных клетках зависает под действием альтернативных сил натяжения между двумя полюсами (рис. 1, а, б), создавая иногда иллюзорность их однополярной ориентации.
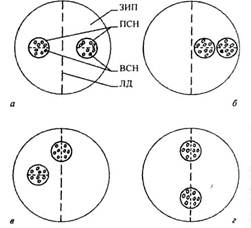
Рис. 4. Варианты расположения точек фиксации пучка связующих нитей дуплицированной центромеры после сегрегации зоны исходного полюса: а, б – расположение по типу эквационного (а) и редукционного (б) делений; в, г – расположение пучка связующих нитей в зоне исходного полюса, способствующего поперечному делению одной (в) и двух (г) хроматид; ЗИП – зона исходного полюса; ЛД – линия деления полюсной зоны; ПСН – пучки связующих нитей дуплицированной центромеры; ВСН – волокна связующих нитей
Полюсной детерминацией характера расхождения хромосом объясняются случаи поперечного деления хроматид и в анафазе II. При расхождении хромосом в анафазе первой их кинетохоры имеют униполярную ориентацию. Соответственно, для перехода на митотический тип деления должна произойти их авто-ориентация. Для хромосомы с двумя сестринскими центромерами данный процесс проходит нормально. Однако когда в него вступает хроматида, имеющая один кинетохор, точки фиксации его связующих нитей часто оказываются в зоне раздела исходного полюса. При полном или частичном совпадении расщепления пучка микротрубочек с линией потенциального раздела центромеры происходит ее поперечное деление. При этом образуются два телоцентрика. По данным литературы [10] и результатам наших исследований, иногда в анафазе II наблюдались случаи, когда хроматида зависала в экваториальной зоне без проявления ее смещения к одному из полюсов. Подобного рода ситуация возникает, если происходит расщепление пучка микротрубочек, соответствующее их продольному распределению по центромере.
В работе J. M. Vega и M. Feldman [10] было показано достоверное влияние гена Ph1 на частоту поперечного деления центромеры унивалента. В наших исследованиях функция данного гена не изучалась. Однако было доказано наличие вмешательства генетического фактора в частоту распределения функций между гомологами при смене их полюсной ориентации [4]. Установленное для моносомной серии соотношение 50 на 50 % в большинстве случаев нарушается в пользу одного из гомологов, когда по одной паре хромосом чаще оказывается задействованной сегрегацией зоны исходного полюса хромосома отцовского родителя, а по другой – материнского. Было показано, что данным обстоятельством достигается возможность избежать синапсис гомеологов. Очевидно, подобного рода вмешательством может объясняться и эффект действия гена Ph1 на misdivision.
Из данных вышеприведенных таблиц (таб. 1, 2) следует, что линии по частоте поперечного деления унивалента существенно различались между собой. Однако слабая корреляционная зависимость между частотой биполярной ориентации унивалента и поперечного деления его центромеры не свидетельствует в пользу центромерной детерминации последнего. Очевидно, частота поперечного деления центромеры унивалента определяется местом положения хромосомы в интерфазном ядре.
Заключение. Результаты анализа характера поведения унивалента в мейозе, проведенного на полной серии моносомных линий мягкой пшеницы, показали, что поперечное деление центромеры унивалента является частным случаем проявления общего механизма перехода хромосом от одного типа деления к другому. При этом была продемонстрирована возможность не только ее продольного расщепления, но и поперечного. Предпринятые статистические расчеты свидетельствуют о наличии определенных элементов случайности в частоте проявления misdivision. На основании ранее опубликованных экспериментальных данных [3, 4] есть основания полагать, что различия между хромосомами по анализируемому мейотическому признаку связаны с их конкретным пространственным положением в интерфазном ядре.
Рецензенты:
Плотникова Людмила Яковлевна, доктор биологических наук, профессор кафедры селекции, генетики и физиологии растений ОмГАУ, г. Омск.
Шаманин Владимир Петрович, доктор сельскохозяйственных наук, профессор кафедры селекции, генетики и физиологии растений ОмГАУ, г. Омск.
Библиографическая ссылка
Жарков Н. А. МЕХАНИЗМ ПОПЕРЕЧНОГО ДЕЛЕНИЯ ЦЕНТРОМЕРЫ УНИВАЛЕНТНОЙ ХРО-МОСОМЫ У TRITICUM AESTIVUM L. // Современные проблемы науки и образования. – 2013. – № 1. ;URL: https://science-education.ru/ru/article/view?id=8120 (дата обращения: 20.04.2024).