



Вопросы профилактики заболеваемости и повышение защитных функций организма ребенка всегда были актуальны [1–3]. Первичным звеном барьера на пути возникновения патологических процессов является клеточная мембрана, в том числе и эритроцитарная [4, 5]. Обязательным условием в сохранении целостности и функциональной активности эритроцитарных мембран (ЭМ) является наличие в их структуре определенного количества фосфолипидов и белков. Кроме того, необходимыми компонентами для стабилизации биологических мембран являются структурообразующие жирорастворимые витамины (ЖРВ): ретинол (РТ) и токоферол (ТКФ) [6, 7], которые локализованы в различных областях фосфолипидов. РТ занимает их поверхностный слой (полярную часть молекул). При этом действие РТ направлено на улучшение обменных процессов. ТКФ располагается в области углеводородных «хвостов» легкоокисляемых фосфолипидов, в зоне их двойных связей. Он способствует восстановлению окисленных форм арахидоновой жирной кислоты, и, следовательно, легкоокисляемые фосфолипиды остаются в структуре биомембран в функционально активном состоянии. Вместе с тем биологически активный ТКФ (восстановленная форма) способствует сохранению структуры и функции высоконенасыщенного липидрастворимого РТ [8].
При изучении нами взаимодействия ЖРВ (РТ и ТКФ), входящих в структуру ЭМ, с липидами, белками и структурированной водой было доказано, что антиокислительные функции альфа-токоферола (α-ТКФ) не всегда связаны с его количественным содержанием в структуре биомембран. В основном они обусловлены качественными характеристиками, то есть биологической активностью [9].
Известно, что при нарушении окислительно-восстановительного баланса в клеточных мембранах накапливаются окисленные формы ТКФ и РТ [8]. В результате эти ЖРВ становятся опасными прооксидантами, так как в таком состоянии быстро запускаются дестабилизационно-деструктивные процессы. Определить грань дефицита или, наоборот, избытка данных витаминов в мембранах в случае нарушения баланса белков, жиров и микроэлементов трудно. И в том и в другом случае окислительные процессы усиливаются и поэтому клинически не имеют различий. Однако для сохранения структуры биомембран и укрепления здоровья ребенка в целом введение в организм этих витаминов в жирорастворимой форме в качестве профилактических мер часто является необходимым. В ранее проведенной нами работе мы исследовали влияние ЖРВ (РТ и ТКФ) на метаболизм структурных компонентов суспензии ЭМ практически здоровых детей (фосфолипидов интегральных белков и структурированной воды) и показали признаки, характеризующие прооксидантные и антиоксидантные свойства ЖРВ (РТ и ТКФ), включенных в структуру ЭМ. При этом нами было установлено их суммарное воздействие на ЭМ [9]. Но вопрос о влиянии на нативную «живую» ЭМ каждого отдельно взятого введенного витамина не был изучен. Научная информация по данному вопросу в анализируемой литературе практически отсутствует. Поэтому целью нашего дальнейшего исследования было изучить в эксперименте методом нагрузочных проб in vitro изолированное и сочетанное воздействие РТ и ТКФ на физико-химические свойства ЭМ практически здоровых детей.
Материал и методы исследования
Были изучены физико-химические характеристики эритроцитарных мембран у 20 практически здоровых детей, проживающих в г. Красноярске, при условиях введения в суспензию ЭМ in vitro липидрастворимых витаминов. В качестве экспериментального материала применялись фармакологические препараты: масляные формы ЖРВ ретинол ацетат (РТА), α-ТКФ и аевит. Исследования проводились в четыре этапа: до введения витаминов, после введения в одну и ту же исходную пробу РТА, α-ТКФ, аевита. В качестве растворителя использовали гексан (1:100000). В 50 мл суспензии мембран эритроцитов добавляли 10 мл раствора витаминов.
Физико-химические свойства эритроцитарных мембран (ЭМ) определялись спектрофлюоресцентным методом с использованием зондов: пирен, 1-анилино-нафталин-8 сульфонат (АНС), 4-диметиламинохалкона (ДМХ), хлортетрациклин гидрохлорид (ХТЦ), нистатин. Данные измерения производились на спектрофлуориметре MPF-4 марки «Хитачи» (Япония). Наряду с этим применялся метод беззондовой флуоресценции: определение степени собственной флуоресценции триптофановых групп белков (длина волны экстинкции − 284 нм, эмиссии − 334 нм) [10] и степени дестабилизации ЭМ по соотношению поляризации (P) и фоновой деполяризации мембраны (dP) (коэффициент оптической лабильности (P/dP) [11]. По эксимеризации пирена определяли степень текучести в 2 зонах его локализации: общего липидного (длина волны экстинкции − 340 нм, эмиссии − 350 нм) и прибелкового (аннулярного) (длина волны экстинкции − 284 нм, эмиссии – 350 нм). Для оценки энерготранспортной функции интегральных белков определяли степень кальцийсвязывающей способности (КСВ) ЭМ c использованием зонда хлортетрациклина гидрохлорид (ХТЦ) (длина волны экстинкции − 380 нм, эмиссии − 400 нм) [10]. С применением отрицательно заряженного зонда (АНС) определяли степень его анизотропии (длина волны испускания – 400 нм, возбуждения – 360 нм) [10]. Определяли степень насыщенности ЭМ структурированной внутримембранной водой двух компонент свечения зонда ДМХ (4-диметиламинохалкона) − длинноволновой (542 нм) и коротковолновой (498 нм). Длина волны возбуждающего света была 427 нм. Для определения количественного содержания воды использовали коэффициент (1/фл ДМХ), так как флуоресценция зонда ДМХ гасится структурированной мембранной водой, окружающей зонд [10]. Степень нарушения липидных кластерных образований гидрофильной области поверхностных фосфолипидов определяли по динамике связывания каналообразующего зонда нистатина (длина волны экстинкции − 320 нм, эмиссии – 460 нм) [10].
Математическую обработку полученных результатов проводили с использованием статистического пакета прикладных программ BIOSTAT, ver. 6.0. (StatSoft Inc. США). Все полученные результаты проверялись на нормальность распределения с помощью критерия Колмогорова–Смирнова. Для количественных признаков использовался непараметрический U-критерий Манна–Уитни при сравнении двух связанных выборок. Изменения считались статистически значимыми при уровне значимости р<0,05. Анализ зависимости признаков проводился с помощью расчета и оценки значимости непараметрического коэффициента корреляции по Спирмену. При значении р<0,05 регрессионная модель адекватно описывает взаимосвязь признаков.
Результаты исследования
Данные об изменениях физико-химических свойств в ЭМ в результате экспериментального введения в одну и ту же исходную пробу практически здоровых детей фармакологических форм препаратов РТА, α-ТКФ и аевита (in vitro) представлены в таблице. Эти изменения носили однозначный характер, несмотря на различную локализацию ЖРВ (ретинола и токоферола) в биомембранах.
Показатели физико-химических изменений в экспериментальных мембранах эритроцитов при нагрузочных пробах с жирорастворимыми витаминами (в условных единицах) (Me 25‰ – 75‰)
|
Показатель |
Исходное состояние |
После введения ретинола ацетат |
После введения α-токоферола |
После введения аевита |
|
1 |
2 |
3 |
4 |
5 |
|
Триптофановые группы белков, фл.ед. |
9,740 (7,850–11,150) |
10,555 (8,9–11,85) |
10,475 (8,5–12,35) Р2-4=0,0990 |
10,8 (8,35–12,65) Р2-5=0,03389 |
|
1 |
2 |
3 |
4 |
5 |
|
Текучесть аннуляр. Ан фосфолипидов, отн. ед. |
0,373 (0,343–0,406) |
0,403 (0,351–0,461) |
0,401 (0,225–0,421) |
0,415 (0,336–0,465) |
|
Текучесть фосфолипи- сло дов общ. билип. слоя, отн. ед. |
0,589 (0,540–0,657) |
0,655 (0,530–0,792) |
0,702 (0,579–0,866) Р2-4=0,0442 |
0,696 (0,602–0,778) Р2-5=0,0155 |
|
Анизотропия зонда АНС, отн. ед. |
0,442 (0,393–0,479) |
0,422 (0,367–0,42) |
0,473 (0,439–0,521) Р2-4=0,0442 |
0,451 (0,403–0,494) |
|
Гидратировонность, отн. ед (1/фл.ДМХ,542) |
0,039 (0,031-0,043) |
0,044 (0,033-0,042) |
0,042 (0,033-0,004) Р2-4=0,0059 |
0,047 (0,031-0,053) Р2-5=0,0339 |
|
Гидратировонность, отн. ед (1/фл.ДМХ, 498) |
0,019 (0,015–0,022) |
0,021 (0,015–0,023) |
0,022 (0,017–0,023) Р2-4=0,0095 |
0,023 (0,017–0,026) Р2-5=0,0139 |
|
Проницаемость по нистатину, отн. ед. |
0,308 (0,169–0,339) |
0,288 (0,215–0,372) |
0,357 (0,225–0,421) |
0,402 (0,285–0,439) Р2-5=0,0665 |
|
Оптическая лабильность (Р/dp), отн. ед. |
26,570 (20,636–34,969) |
27,245 (21,846–32,300) |
26,227 (22,705–29,485) |
26,325 (19,756–30,249) |
|
Кальцийсвязывающая щая способ. м.э., отн. ед. |
9,927 (4,889–16,403) |
9,565 (3,497–13,462) |
9,836 (3,841–11,075) |
11,058 (3,70–14,77) |
Установлено, что в пробах после введения в них (in vitro) изолированных РТА и α-ТКФ и сочетанного препарата аевит были обнаружены значительные изменения их физико-химических характеристик. Эти трансформации произошли как в гидрофобной области фосфолипидов общего билипидного, так и в поверхностном белково-липидном слое ЭМ за счет роста белкового фактора (триптофанилов) и изменения степени подвижности фосфолипидов. Выявлено повышение флуоресценции показателя триптофанилов, наиболее выраженное в пробе с аевитом (р=0,0339). Вместе с тем был отмечен рост гидратированности экспериментальных ЭМ с добавленными в исходную пробу витаминами (α-ТКФ и аевит), причем как в гидрофильной (542 нм), так и в гидрофобной (498 нм) компоненте (таблица). Это повышение составило для α-ТКФ в гидрофильной зоне фосфолипидов 7,1% (р=0,0059), для аевита 17,0% (р=0,0339). В гидрофобной области фосфолипидов для α-ТКФ 13,6% (р=0,0095), для аевита 17,4% (р=0,0139). Кроме того, отмечено повышение текучести углеводородов фосфолипидов общего билипидного слоя после введения РТА на 11,42%, α-ТКФ − на 16,1% (р=0,0442) и аевита − на 15,4% (р=0,0155) по сравнению с исходной пробой (таблица). В эксперименте с аевитом выявлена тенденция к повышению степени проницаемости фосфолипидов ЭМ по нистатину в их гидрофильной области на 23,4% (р=0,0665). В эксперименте с α-ТКФ установлено повышение показателя степени анизотропии зонда АНС, встроенного в структуру фосфолипидов, в месте его контакта с интегральным белком (р=0,0410) (таблица), что соответствовало снижению подвижности гидрофильных участков фосфолипидов в данной области ЭМ.
Полученные результаты в основном подтверждают те свойства, которые описаны ранее на модельных мембранах [8]. При проведении сравнительного корреляционного анализа между исследуемыми показателями физико-химических свойств в исходной пробе и витаминизированных пробах (in vitro) были выявлены особенности молекулярных взаимодействий, характерных только для исходной пробы и отсутствующих в пробах с добавленными витаминами (рис. 1).
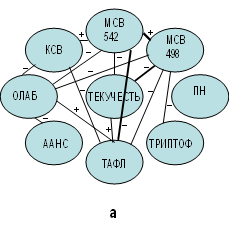
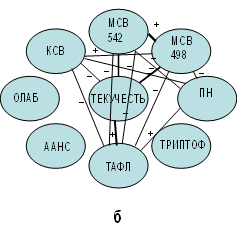
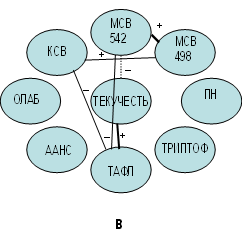
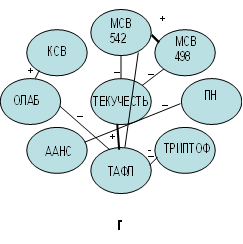
Рис. 1. Корреляционные взаимосвязи между показателями физико-химических параметров
эритроцитарных мембран, отражающих молекулярные изменения в результате
проведенного эксперимента: а – исходная проба; б – эксперимент с добавленным ретинолом ацетат; в – эксперимент с добавленным альфа-токоферолом; г – эксперимент
с добавленным аевитом
Примечание. ТЕКУЧЕСТЬ – текучесть углеводородов фосфолипидов билипидного слоя;
ТАФЛ – текучесть углеводородов аннулярного слоя фосфолипидов;
МСВ 542 – мембраносвязанная вода гидрофильная компонента;
МСВ 498 – мембраносвязанная вода гидрофобная компонента;
КСВ – кальцийсвязывающая способность;
ОЛАБ – оптическая лабильность;
ААНС – анизотропия зонда АНС;
ТРИПТОФ – триптофанилы;
ПН – проницаемость мембран эритроцитов по нистатину.
Эти особенности связаны с молекулярными взаимодействиями мембраносвязанной воды (МСВ) с исследуемыми показателями. Во-первых, в исходной пробе количество корреляционных взаимодействий МСВ значительно превосходит экспериментальные пробы с добавленными в них витаминами (рис. 1а). Во-вторых, только в исходной пробе нами установлена взаимосвязь показателя триптофанилов (ТРИПТОФ) с гидрофобной компонентой МСВ (498 нм) (r=−0,4600, p=0,0413) (рис. 1а). Данная взаимосвязь указывает на то, что координирующее центры углеводородной зоны фосфолипидов, окружающих и активизирующих интегральные белки образованы с помощью МСВ. Это свидетельствует о ее стабилизирующих свойствах, сохраняющих структуру транспортных белков [7], тогда как выявленная, характерная только для исходной пробы прямая корреляционная взаимосвязь гидрофильной компоненты воды (МСВ) (542 нм) с показателем, характеризующим функции данных белков (кальцийсвязывающая способность) КСВ (r=0,4597, p=0,0414) (рис. 1а), свидетельствует об участии гидрофильной компоненты МСВ в регуляции энергозависимого транспорта МЭ. Данный факт может указывать на участие МСВ в ионнотранспортной функции, что было отмечено нами ранее [12]. Вместе с тем установлено, что в исходной пробе суспензии ЭМ МСВ структурирует фосфолипиды как общего, так и прибелкового слоев. Данное суждение основано на том, что МСВ имеет отрицательные взаимосвязи с углеводородами фосфолипидов общего билипидного слоя с гидрофильной (542 нм) (r=−0,627, р=0,0031) в зоне «головки» фосфолипидов и гидрофобной (498 нм) (r =−0,5527, р=0,0115) в области углеводородов (рис. 1а). При отсутствии значимых различий в показателе текучести аннулярных фосфолипидов (ТАФЛ), отвечающих за активацию транспортных белков, отмечена его обратная взаимосвязь с МСВ (542 нм) (r=−0,7539, р=0,0001) и МСВ (498 нм) (r=−0,5974, р=0,0054). Следовательно, нами доказано, что в нативной ЭМ исходной пробы фосфолипиды и, соответственно, интегральные белки окружены структурированными молекулами воды (аквасферами), сохраняющими их стабильность и функции [7]. Подтверждением этому являются выявленные также только в исходной пробе обратные корреляционные взаимосвязи МСВ (двух ее компонент) с показателем оптической лабильности (ОЛАБ) (542 нм) (r=−0,4816, p=0,0316) и (498 нм) (r=−0,5648, p=0,0095). Этот показатель косвенно характеризует процессы дестабилизации (рис. 1а), что подчеркивает, что МСВ в ЭМ исходной пробы способствует структурированию фосфолипидов и интегральных белков [7] и обеспечивает их нормальное функционирование.
Выявленные особенности молекулярных взаимодействий в исходной пробе были характерны для мембран эритроцитов здоровых детей с достаточным содержанием в структуре ЭМ ЖРВ (РТ и ТКФ) со сбалансированной окислительно-восстановительной системой. Препарат РТА фармакологической формы, введенный в эритроцитарную суспензию исходной пробы, способствовал модификации ее молекулярной структуры. Корреляционный анализ показал, что в экспериментальной пробе с РТА ведущее место в регуляции функции ЭМ занимает показатель, характеризующий проницаемость поверхностного фосфолипидного слоя, - каналообразующий зонд нистатин. Он локализуется в липидной фазе мембран и образует поры, через которые происходит утечка метаболитов из цитоплазмы [10]. Наличие множества корреляционных взаимодействий показателя проницаемости (ПН) МЭ с другими физико-химическими показателями (рис. 1б) при отсутствии физико-химических различий с исходной пробой свидетельствует скорее о качественных изменениях канальной системы поверхностного фосфолипидного слоя МЭ. Установленные обратные корреляционные взаимодействия, ПН и МСВ, в двух компонентах свечения ДМХ гидрофильной (542 нм) (r=−0,4612, p=0,0407) и гидрофобной (498 нм) (r=−0,4716, p=0,0358) могут свидетельствовать о том, что при условии повышения ионной проницаемости (количества ионных каналов) МЭ структурообразование мембранной воды будет снижаться. По нашей гипотезе это может быть связано с взаимопревращением неустойчивых структурированных водных каналов в поры в момент их расширения в результате повышения ПОЛ или с различным состоянием порообразования [7]. Если проследить дальнейшую цепочку корреляционных взаимодействий ПН, то выявится неблагоприятное влияние ионной проницаемости (ПН) суспензии МЭ с добавленным витамином РТА на текучесть углеводородов фосфолипидов, локализованных в прибелковой области (ТАФЛ) (r=0,5130, p=0,021) (рис. 1б). При условии повышения ионной проницаемости однозначно будет повышаться текучесть фосфолипидов. На первый взгляд это является позитивным фактором. Однако условное повышение текучести аннулярных фосфолипидов (ТАФЛ) и повышение ионной проницаемости (ПН) будут способствовать снижению энерготранспортной функции (КСВ) ЭМ. Данное суждение основано на полученных обратных корреляционных взаимосвязях показателя кальцийсвязывающей способности (КСВ) с показателями ПН (r=−0,5175, p=0,0194) и ТАФЛ (r=−0,5077, p=0,0223) (рис. 1б). Это в совокупности свидетельствует о том, что в случае повышения проницаемости фосфолипидного слоя и текучести прибелковых фосфолипидов будет снижаться энергозависимый транспорт через ЭМ. Следовательно, повышение текучести обусловлено дестабилизационными процессами в суспензии с добавленным витамином А. Происходят ослабление и потеря связи белковых компонентов ЭМ (триптофанилов) (ТРИПТОФ) с МСВ. Результатом этого является отсутствие их взаимодействия, что способствует повышению дестабилизационных процессов.
При добавлении в исходную суспензию ЭМ α-ТКФ выявлено, что молекулярные изменения носили по сравнению с предыдущей анализируемой пробой иной характер (рис. 1б,в). Мы отметили значительно меньшее число корреляционных взаимосвязей в пробе с добавленным α-ТКФ. Установлено, что повышение в данном опыте содержания МСВ по сравнению с исходной пробой способствует активации кальцийсвязывающей способности (КСВ) МЭ. Об этом свидетельствует прямая взаимосвязь МСВ (498 нм) и КСВ (r= 0,4754, p=0,0341). Повышению уровня МСВ гидрофильной компоненты (542 нм) будет способствовать снижение степени текучести фосфолипидов прибелковой области МЭ (ТАФЛ), о чем свидетельствует обратная корреляционная взаимосвязь (r=−0,5498, p=0,0120) (рис. 1в), которая указывает на структуросохраняющие способности кластеров МСВ, расположенных вокруг координирующих центров «головок» фосфолипидов в прибелковой области. Следует отметить, что в экспериментальной пробе с добавленным α-ТКФ зависимость текучести углеводородной части фосфолипидов общего билипидного слоя ЭМ (ТЕКУЧЕСТЬ) с МСВ не выявлена (рис. 1в), что свидетельствует о вытеснении или разрушении слабых ковалентных связей водных кластеров вокруг углеводородов фосфолипидов общего билипидного слоя ЭМ [7]. Это соответствует свойствам липидрастворимого витамина α-ТКФ, а именно локализации его в углеводородной зоне легкоокисляемых фосфолипидов [7, 8]. Фармакологический препарат α-ТКФ, введенный в исходную суспензию, аналогично естественному способствует сохранению структуры ЭМ. Однако при этом нарушается тонкая регуляция метаболизма, которая осуществляется с помощью МСВ, находящейся во взаимосвязи с интегральными белками и фосфолипидами общего билипидного слоя. В данной части эксперимента такая взаимосвязь отсутствует.
Следует отметить, что воздействие на ЭМ ЖРВ в сочетанной форме (аевит) не оказывало значительного влияния на состояние МСВ. Вместе с тем сравнительный анализ корреляционных взаимосвязей до и после введения витамина аевита в исходную суспензию ЭМ свидетельствовал, что степень связывания показателей МСВ с текучестью фосфолипидов уменьшалась по сравнению с исходной суспензией. Так, выявлено, что корреляционная взаимосвязь показателя степени текучести ЭМ общего билипидного слоя (ТЕКУЧЕСТЬ) с гидрофобной компонентой воды (498 нм) с добавленным аевитом была менее тесная, чем в исходной пробе (r=−0,4635, p=0,0395 против r=−0,5527, p=0,0115 в исходной пробе), с гидрофильной компонентой воды (542 нм) (r=−0,5719, p=0,0084 против r=−0,6267, p=0,0031 в исходной суспензии). Ослабление связей между вышеуказанными показателями может свидетельствовать о некотором «разрыхлении» слабых водородных соединений в структурированной мембранной воде в зоне структурообразующих фосфолипидов. Это, по нашему мнению, является одной из причин повышения степени текучести в зоне структурных фосфолипидов общего билипидного слоя ЭМ (таблица). Она может быть инициирована структурными дефектами в зоне ненасыщенных связей фосфолипидов в результате изменения свойств структурированной воды [7, 12]. По всей видимости, данные процессы повлекли за собой повышение ионной проницаемости ЭМ (ПН) (таблица). Это свидетельствует о дестабилизационных изменениях в МЭ, вызванных ослаблением связей МСВ с фосфолипидами как между собой, так и между интегральными белками, на что указывает прямая взаимосвязь показателей ПН и ААНС (r=0,5263, p=0,0171). Вместе с тем следует отметить характерные для экспериментальной пробы с добавленным витамином аевитом обратные корреляционные связи между показателями текучести фосфолипидов прибелковой области (ТАФЛ) и показателями уровня интегральных белков (ТРИПТОФ) (r=−0,5246, p=0,0175), а также степени дестабилизации (ОЛАБ) (r=−0,5338, p=0,0153) (рис. 1г). Обратный характер этих взаимосвязей указывает на тот факт, что при условии повышения текучести углеводородов фосфолипидов в прибелковой области (ТАФЛ) будет снижаться уровень интегральных белков (триптофанилов), что является неблагоприятным признаком, так как это может быть обусловлено ПОЛ. Что же касается общей стабильности (ОЛАБ), то на данном этапе фосфолипиды аннулярной прибелковой области способствуют сохранению стабильности ЭМ. Это в свою очередь будет содействовать активации энерготранспортной функции интегральных белков ЭМ с аевитом, о чем свидетельствует прямая взаимосвязь показателей ОЛАБ и КСВ (r=0,5459, p=0,0128). Следовательно, введение сочетанной формы ЖРВ аевита в суспензию исходных ЭМ более благоприятно для них, чем введение изолированных форм. Однако и в данном случае, так же как в предыдущих эпизодах, имеются негативные признаки. Это, с одной стороны, разрыхление водных кластеров, а именно отсутствие взаимосвязей МСВ с ОЛАБ и ТРИПТОФ, с другой – наличие обратной взаимосвязи между показателями ТАФЛ и ТРИПТОФ.
Таким образом, нами установлено, что при достаточной антиокислительной активности ЭМ структурные компоненты (фосфолипиды, белки и МСВ) находятся в тесной взаимосвязи между собой. Это способствует сохранению структуры и функции ЭМ у исследуемых детей. Жирорастворимые витамины РТ и ТКФ, входящие в структуру ЭМ, в данном случае не нарушают кластерные образования МСВ, а действуют согласованно с ними. Стабильность структуры суспензии ЭМ у здоровых детей (фосфолипиды и белки) поддерживается с помощью структурированной в кластеры мембранной воды ЖРВ (РТ и ТКФ). Все метаболические процессы обмена функционируют сообразно физиологическим нормам. При этом МСВ регулирует все процессы обмена, в том числе она осуществляет так называемую тонкую регуляцию, когда воздействие на рецепторные белки происходит через молекулы воды [13]. Вместе с тем МСВ не только участвует в каналообразовании в фосфолипидном слое, но и образует канальную систему в белковой составляющей и таким способом оказывает позитивное влияние на энерготранспортную функцию ЭМ. Именно это обеспечивает биомембране нормальное функционирование и свидетельствует о достаточной биологической активности структурных внутримембранных ЖРВ. Потеря и ослабление связей основных структурообразующих компонентов биомембран (белков и фосфолипидов) со структурированной водой инициируют первичные дестабилизационно-деструктивные процессы [7, 12].
Введение препаратов жирорастворимых витаминов фармакологического производства в любой форме (изолированной или сочетанной) способствовало ослаблению и разрыву слабых водородных связей структурированной воды с основными структурными компонентами биомембран (белками и фосфолипидами), так как эти витамины по своей сути являются ксенобиотиками, то есть чужеродными веществами, которые разрушают эти связи и способствуют дестабилизационным изменениям. Эти процессы неблагоприятны для функционирования ЭМ, особенно в условиях роста и развития ребенка.
В результате проведенного экспериментального исследования получены новые данные о молекулярных изменениях в нативных «живых» ЭМ у практически здоровых детей при добавлении в их ЭМ (in vitro) препаратов фармакологического производства ЖРВ (РТА, α-ТКФ и аевита) для профилактики гиповитаминозов. Установленные нами закономерности указывают на необходимость разработки новых подходов как к лечению с помощью данных витаминов, так и к профилактическим мероприятиям, особенно в раннем детстве.
Библиографическая ссылка
Колодяжная Т.А. ИЗУЧЕНИЕ В ЭКСПЕРИМЕНТЕ МЕТОДОМ НАГРУЗОЧНЫХ ПРОБ (IN VITRO) СОЧЕТАННОГО И ИЗОЛИРОВАННОГО ДЕЙСТВИЯ РЕТИНОЛА АЦЕТАТ И Α-ТОКОФЕРОЛА НА ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА МЕМБРАН ЭРИТРОЦИТОВ // Современные проблемы науки и образования. – 2018. – № 6. ;URL: https://science-education.ru/ru/article/view?id=28200 (дата обращения: 19.04.2024).