



Галофитные и гликофитные виды растений имеют некоторые общие механизмы устойчивости к засолению, прежде всего это стресс-индуцированное накопление осмопротекторов и активизация антиоксидантной защиты. В функционировании их защитных систем важнейшее значение отводится пролину, защищающему белково-липидные комплексы путем обезвреживания гидроксильных радикалов и других активных форм кислорода, повреждающих биомембраны. Пролин вовлекается в сложную регуляцию генов различных изоформ супероксиддисмутазы. При засолении почв растения испытывают водный дефицит; пролин – один из самых распространенных осмолитов, поддерживающих осмотический потенциал клетки [2; 5-7; 9]. Кроме того, J.H. Venekamp (1989) [12] полагает, что образование пролина в условиях стресса является способом регуляции рН цитозоля.
Во многих работах выявлена связь между накоплением пролина и устойчивостью растений к стрессорам. Однако зарегистрированы и противоположные эффекты. Неоднозначность такой связи может быть обусловлена как методическими причинами (различная сила стрессовых воздействий в разных экспериментах), так и сложным взаимодействием пролина с другими стресспротекторными системами, в частности с ферментативной антиоксидантной. Сравнительные исследования такого взаимодействия у сортов и видов культурных растений, отличающихся по устойчивости к стрессорам, могут быть полезны как для разработки адекватных подходов к оценке устойчивости растений, так и для более глубокого понимания стресспротекторных функций пролина [8].
В почвах накопление легкорастворимых солей при определенном их составе может сопровождаться увеличением рН до 9-10 единиц. В г. Соликамске Пермского края на территории солеотвалов реакция почвенной среды достигает щелочных значений (8,5-8,7 рН). На расстоянии 1-5 м от солеотвала в корнеобитаемом слое почв общее содержание водорастворимых солей составляло от 0,3 до 2,2%, на расстоянии 5-100 м – от 0,01 до 0,3% [4]. Сочетание засоленности и щелочности требует от растений из зоны воздействия солеотвалов эффективных механизмов адаптации. В современной физиологии растений практически отсутствуют сведения о совместном воздействии на высшие растения засоления и щелочности корневой среды. Таким образом, исследование механизмов устойчивости растений к совместному воздействию засоления и щелочности, в том числе с участием универсального осмопротектора – пролина, является актуальным.
Материалы и методы исследований
Цель работы – установить изменение в содержании пролина в связи с адаптацией растений к совместному воздействию NaCl-засоления и щелочности корневой среды.
В корнеобитаемом слое почв реакцию среды измеряли на иономере, водорастворимый Na+ – методом пламенной фотометрии; содержание свободных ионов Cl– – меркурометрическим методом. Содержание пролина и солей исследовали у видов растений (Triglochin maritima L., Tussilago farfara L., Plantago media L.), обитающих в зоне воздействия солеотвалов. Модельные опыты по устойчивости проведены с овсом посевным Avena sativa L. и кресс-салатом Lepidium sativum L. Растения выращивали на вермикулите с раствором Кнопа в течение 5-7 дней, затем в вермикулит добавляли растворы едкого натра и хлористого натрия для формирования солевой и щелочной среды. Содержание пролина и свободных ионов изучали через 10-12 часов после стресса.
Пролин в листьях и подземных органах определили по Bates et al. В водной вытяжке из органов растений содержание свободных ионов Na+ измерили методом пламенной фотометрии; содержание свободных ионов Cl– – меркурометрическим методом. Сбор растений у солеотвалов был проведен в 10-12 повторностях. В модельных опытах повторность определений – 3-кратная. Замеры высоты и массы растений в 25-кратной повторности провели через сутки после воздействия стресса. Достоверность различий между вариантами устанавливали дисперсионным методом по критерию Фишера и уровню значимости нулевой гипотезы; взаимосвязи между показателями состояния растений оценивали методом корреляционного и регрессионного анализов.
Результаты и обсуждение
У солеотвалов, в местах обитания триостренника и мать-и-мачехи, почвы характеризовались засоленностью и щелочностью; а подорожник средний произрастал при минимальном количестве солей, слабокислой и нейтральной реакции почвенной среды (табл. 1).
Таблица 1
Содержание водорастворимых ионов (мг-экв/100 г) и рН в корнеобитаемом слое местообитаний растений у солеотвалов
|
Вид |
рН |
Na+ |
Cl- |
|
Триостренник приморский |
6,9 – 8,6 |
1,7 – 36,5 |
0,1 – 34,5 |
|
Мать-и-мачеха обыкновенная |
6,9 – 8,7 |
0,7 – 3,0 |
0,3 – 3,7 |
|
Подорожник средний |
5,6 – 7,3 |
0,1 – 1,0 |
0,1 – 1,1 |
Наибольшее количество водорастворимых ионов как в листьях, так и в подземных органах характерно для солеустойчивого триостренника приморского, относящегося к видам с высоким конститутивным уровнем пролина (табл. 2).
Таблица 2
Содержание свободных ионов и пролина в растениях, произрастающих в зоне воздействия солеотвалов, мг/100 г сухой массы
|
Вид |
Органы |
Na+ |
Cl- |
Пролин |
|
Триостренник приморский |
Листья |
4556 ± 86 |
4922 ± 107 |
2118 ± 199 |
|
Подземные органы |
1453 ± 250 |
1470 ± 188 |
847 ± 242 |
|
|
Мать-и-мачеха обыкновенная |
Листья |
877 ± 74 |
1213 ± 49 |
60 ± 7 |
|
Подземные органы |
929 ± 80 |
978 ± 59 |
119 ± 13 |
|
|
Подорожник средний |
Листья |
440 ± 49 |
2382 ± 142 |
25 ± 6 |
|
Подземные органы |
637 ± 55 |
994 ± 78 |
21 ± 1 |
У триостренника проявилось стресс-индуцированное накопление пролина, т.к. установлена прямая зависимость между пролином и количеством свободных ионов Cl– в листьях (мг/100 г сухой массы): у = 1,13*x–3328; коэффициент корреляции R=0,73; коэффициент детерминации R2=0,53; критерий Фишера F=6,9; уровень значимости нулевой гипотезы р=0,0008.
Для мать-и-мачехи характерно среднее количество солей и пролина, уровень пролина повышен в корневищах по сравнению с листьями. Одновременно не установлена связь между содержанием этого осмопротектора и засоляющих ионов в органах растения. У подорожника с минимальным накоплением свободных ионов выявлен наименьший пул пролина в листьях и подземных органах; у него также отсутствует связь между накоплением этих соединений.
Овес выращивали на вариантах разного засоления NaCl, при 7 и 9 рН. С увеличением концентрации хлорида натрия (от 0 до 2%) уровень накопления свободных ионов Cl- в листьях овса возрастал – от 2400 до 7600 мг/100 г, а свободных ионов Na+ – от 500 до 1500 мг/100 г сухой массы. Содержание Na+ существенно ниже, чем Cl-, что характерно для растений, обитающих в условиях природного засоления-осолонцевания почв [10], а также техногенного засоления почвоподобных образований [4].
В листьях и корнях овса посевного на всех вариантах засоления корневой среды отмечено более высокое содержание пролина относительно контроля (рис. 1). При этом количество пролина достоверно возрастало от контрольного уровня до варианта со средним засолением (0,6% NaCl), но при более высоком засолении (1-2%) его уровень падал, вероятно, из-за угнетения растений.
При щелочности корневой среды (9 рН) на фоне засоления прослежена некоторая тенденция к накоплению пролина в листьях и корнях (рис. 1, 2). Двухфакторный дисперсионный анализ подтвердил достоверное влияние высокой щелочности при разном уровне засоления, различия в накоплении пролина в корнях относительно вариантов отдельного засоления (F=4,26; р=0,0002).
Таким образом, подтвердилось участие пролина в формировании устойчивости овса посевного, т.к. засоление корневой среды сопровождалось его аккумуляцией в листьях; однако при прогрессирующем засолении и угнетении растений его количество снижалось. При совместном воздействии факторов тенденция к накоплению пролина в листьях и корнях овса на фоне щелочности оказалась математически не достоверной при данном объеме выборки.
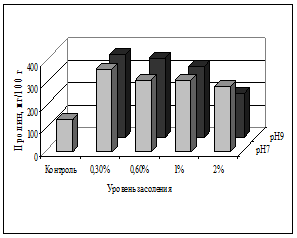
Рис. 1. Содержание пролина в листьях овса посевного, мг/100 г сухой массы
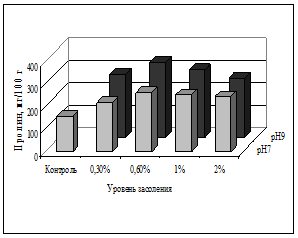
Рис. 2. Содержание пролина в корнях овса посевного, мг/100 г сухой массы
Кресс-салат выращивали при разной щелочности (6-10 рН) и одном уровне засоления корневой среды (0,6% NaCl); ранее проводимые исследования показали, что кресс-салат способен адаптироваться к этой концентрации [3]. Растения в листьях аккумулировали свободные ионы натрия и хлора: Na+ от 300 мг/100 г – на контроле и до 800 мг/100 г – при засолении, Cl- от 1000 мг/100 г и до 1700 мг/100 г соответственно (рис. 3, 4). Накопление ионов Cl- было максимальным на фоне наибольшей щелочности, возможно, при этом происходило нарушение барьерных функций мембран. По количеству ионов Na+ в кресс-салате достоверно отличались варианты с внесением хлоридов натрия от вариантов с отдельной щелочностью. Содержание Na+ в растениях заметно ниже, чем Cl-, преимущественное накопление хлоридов растениями отмечено в условиях засоления-осолонцевания почв [4; 10].
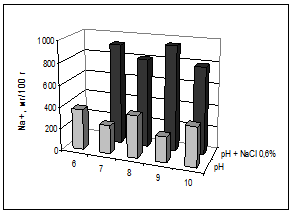
Рис. 3. Содержание свободных ионов натрия в листьях кресс-салата, мг/100 г сухой массы
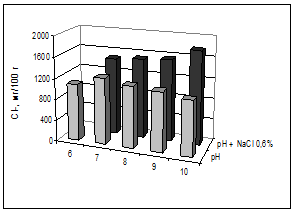
Рис. 4. Содержание свободных ионов хлора в листьях кресс-салата, мг/100 г сухой массы
Кресс-салат отличался от овса более высоким конститутивным уровнем пролина (рис. 5). Содержание пролина (мг/г) в листьях изменялось прямо пропорционально количеству свободных ионов Cl- (мг/г): y=0,82+0,51 x, R=0,72, R2=0,52, F=7,5, p=0,0003. Значение пролина в солеустойчивости кресс-салата подтвердила сильная отрицательная зависимость массы одного растения (y) от содержания свободных ионов хлора (х1, мг/г) и пролина (х2, мг/г): y=40–0,53x1–0,89x2, R= – 0,82, R2=0,68, F=6,3, р=0.034. Следовательно, засоление привело к накоплению в листьях ионов Cl- и пролина, выполняющего осмопротекторную функцию, одновременно снижалась масса растений.
У кресс-салата пролин, по-видимому, участвует в адаптации к щелочности корневой среды. Особенно повышалось количество пролина в листьях на фоне засоления и усиления щелочности от 6 до 9 рН (рис. 5). Установлена сильная прямая зависимость между пролином (мг/г) и реакцией корневой среды (в интервале 6-9 рН): y=1,61x–3,36; R=0,97; R2=0,93; F=29,5; p=0,0009. На вариантах с наибольшей щелочностью и при совместном воздействии засоления и щелочности (NaCl 0,6%+рН 10) отмечено относительное понижение количества пролина, по-видимому, это связано с общим угнетением растений при токсичности корневой среды.
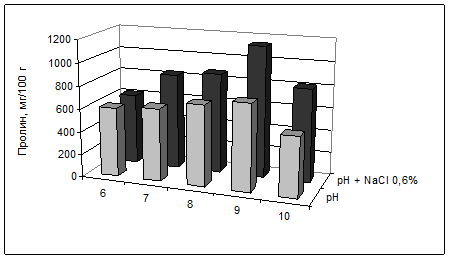
Рис. 5. Содержание пролина в листьях кресс-салата, мг/100 г сухой массы
Через сутки после воздействия стресса было изучено состояние кресс-салата; дисперсионный анализ данных показал достоверное влияние солевых растворов на массу растений в соответствии с критерием Фишера F=37,8 и уровнем значимости нулевой гипотезы р=0,0002. На фоне разных показателей рН засоление понижало среднюю массу одного растения на 4-6 мг, или на 12-21%, относительно варианта без воздействия соли. Солевой стресс также достоверно снизил высоту растений от 5 до 24% по вариантам опыта (F=9,69; р=0,0015). Известно, что на стадии адаптационного процесса мобилизуются защитные системы растений, обеспечивающие избирательное накопление ионов за счет процессов активного транспорта. Происходит дифференцированная экспрессия генов водного дефицита, которые кодируют синтез осмопротекторов разной природы воздействия [1; 2; 11]. Все это энергозависимые процессы, поэтому остается меньше ресурсов на увеличение массы и высоты растений.
Математическая обработка данных не выявила достоверного влияния щелочности на высоту и массу растений, это может быть связано как с кратковременностью воздействия этого фактора, так и с эффективностью адаптационных механизмов.
Заключение
В зоне воздействия солеотвалов более солеустойчивые растения – мать-и-мачеха и особенно триостренник, произрастающие на засоленных и щелочных почвах, характеризовались повышенным количеством пролина в листьях и надземных органах. В модельных опытах у растений с разным конститутивным уровнем пролина (овес посевной и кресс-салат) стресс-индуцированная реакция на засоление корневой среды сопровождалась аккумуляцией этого осмопротектора; однако при наибольшем засолении и угнетении растений прослежено относительное уменьшение его содержания. Пролин, по-видимому, участвует в адаптации кресс-салата к щелочности, т.к. дополнительное его накопление прослежено при увеличении рН от 6 до 9 единиц. У овса посевного на фоне засоления и воздействия высокой щелочи прослежена тенденция к аккумуляции пролина, достоверность которой в корнях овса подтверждена математически.
Актуальность проводимых исследований связана с необходимостью восстановления ландшафтов, испытывающих техногенное засоление и ощелачивание. Опираясь на полученные результаты полевых и модельных работ, можно утверждать, что при оценке фиторемедиационного потенциала растений следует учитывать конститутивный уровень пролина.
Библиографическая ссылка
Четина О.А., Еремченко О.З., Боталова К.И., Середа А.М. ИЗМЕНЕНИЕ В СОДЕРЖАНИИ ПРОЛИНА В РАСТЕНИЯХ ПРИ ВОЗДЕЙСТВИИ NACL-ЗАСОЛЕНИЯ И ЩЕЛОЧНОСТИ КОРНЕВОЙ СРЕДЫ // Современные проблемы науки и образования. – 2016. – № 6. ;URL: https://science-education.ru/ru/article/view?id=26034 (дата обращения: 25.04.2024).