



Цель исследований состояла в выявлении многообразия межпопуляционных взаимоотношений видов в агроценозах смородины черной и изучении влияния антагониста Pseudomonas fluorescens на параметры паразитической активности и размер экологической ниши S.ribis.
Методика исследований
Исследования проводили в 2006-2013 гг. на Новосибирской ЗПЯОС им. Мичурина и частном селекционно-технологическом питомнике Соловьевой А.Е., расположенных в северной лесостепной зоне Новосибирской области. Годы исследований характеризовались различными погодными условиями. Из восьми лет исследований 4 года (2006, 2007, 2009, 2013) были влажными (ГТК>1) и 3 (2010, 2011, 2012) - засушливыми (ГТК<1), условия 2008 года были близки к средним многолетним показателям (ГТК=1,01). Учеты количественных параметров паразитической активности S. ribis и G. ribis проводили каждые 5-7 дней [1]. Степень реализации экологической ниши оценивали по шкале-трафарету [3]. Межпопуляционные взаимоотношения оценивали, используя коэффициент общности (Жаккара), который является показателем количества общих признаков в сравниваемых фитоценозах и других таксонах. Численность эпифитов определяли методом смыва на КАА, МПА, ЧА. Изучение антагонистической активности Ps. fluorescens против S. ribis проводили путем нанесения бактериальной суспензии смеси штаммов с концентрацией 2,5×1010 кл./мл на опавшие и вегетирующие листья смородины в норме 50 г/га. Статистическая обработка экспериментальных данных проведена методом дисперсионного анализа с использованием пакета прикладных программ SNEDECOR [2].
Результаты исследований и их обсуждение
Таксономический состав биоценоза был представлен 12 видами и 2 группами организмов, среди которых к деструктивной биоте относились 10 видов. Наибольшую распространенность в годы исследований имели специализированные фитопатогены (Septoria ribis, Gloeosporium ribis), фитофаги (Cecidophyes ribis, Cryptomyzus ribis) и сапротрофные микроорганизмы филлопланы и ризосферы.
Указанные организмы вступали между собой в следующие формы межпопуляционных отношений: паразитизм, конкуренция, антагонизм, нейтрализм, протокооперация и неспецифическое угнетение (рис. 1).
В агроценозах смородины конкурентная форма взаимоотношений выявлена между фитопатогенами и эпифитными микроорганизмами (грибы, бактерии, актиномицеты) на листьях смородины. Наиболее сильное перекрытие экологических ниш отмечено у грибов S. ribis и G. ribis и эпифитами, коэффициент общности Жаккара составил 0,5-1,0.
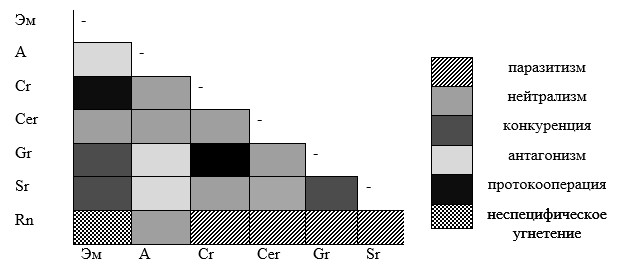
Рис. 1. Формы межпопуляционных отношений в агроценозах черной смородины
Примечание: Rn - Ribes nigrum; Sr - Septoria ribis; Gr - Gloeosporium ribis; Cer - Cecidophyes ribis; Cr- Cryptomyzus ribis; A - антагонисты; Эм - эпифитная микрофлора
Паразитическая форма взаимоотношений выявлена между смородиной и фитопатогенами (S. ribis, G. ribis), смородиной и фитофагами (Cecidophyes ribis, Cryptomyzus ribis). Антагонистические взаимоотношения имели место между фитопатогенными микромицетами и сапротрофными микроорганизмами ризосферы (Pseudomonas fluorescens, Streptomyces spp.).
Нейтральная форма взаимоотношений часто формируется между видами, занимающими разные экологические ниши: смородинный почковый клещ (Cecidophyes ribis) и фитопатогенные микромицеты, антагонисты и фитофаги. Коэффициент общности составил 0-0,5. При нейтрализме наблюдается полное несовпадение или незначительное перекрывание экологических ниш.
Взаимоотношения по типу протокооперация в агроценозах смородины формировалось между листовой галловой тлей (Cryptomyzus ribis L.) и эпифитной микрофлорой, Cr. ribis и G. ribis . Перекрытие экологических ниш Cr. ribis, эпифитов и G. ribis составляло от 80 до 100%.
Между смородиной черной и эпифитными организмами (грибы, бактерии, актиномицеты) на листьях формировались отношения по типу неспецифического угнетения, особенно на устойчивых сортах (табл. 1).
Таблица 1
Численность микрофлоры на листьях смородины, 103/см2 листа
|
Сорт |
Грибы |
Бактерии |
Актиномицеты |
Всего |
|
Памяти Потапенко |
50,8 |
11,2 |
2,4 |
64,4 |
|
Плотнокистная |
12,4 |
5,6 |
0,8 |
18,8 |
|
Дикорастущая смородина |
6,2 |
2,2 |
0,4 |
8,8 |
|
НСР05 |
0,45 |
0,37 |
0,36 |
0,45 |
Данные таблицы свидетельствуют о том, что одним из факторов неспецифической устойчивости сортов смородины черной к фитопатогенам являются фитонцидные выделения листьев [4]. Восприимчивый сорт Памяти Потапенко имел на листьях в 3,4 раза большую численность микроорганизмов по сравнению с высокоустойчивым сортом Плотнокистная и в 7,3 раза по сравнению с дикорастущей смородиной. Дикорастущая смородина имела самую низкую численность всех групп эпифитных микроорганизмов на листьях, что косвенно свидетельствует о максимальной силе ее природных защитных барьеров, по сравнению с измененными селекциями сортовыми генотипами. Высокоустойчивый сорт Плотнокистная снижал численность грибов в 4 раза, бактерий - в 2 раза, актиномицетов - в 3 раза по сравнению с восприимчивым сортом Памяти Потапенко [5]. При добавлении в питательные среды (КАА, МПА, ЧА) отвара из листьев устойчивых сортов наблюдалось снижение скорости роста эпифитной микрофлоры, в частности бактерий и грибов.
Наблюдения за развитием комплекса фитопатогенов на сортах смородины черной позволили сделать вывод, что смородина проявляла неспецифическую комплексную устойчивость к болезням, а патогенные микромицеты имели биотические отношения конкурентного типа. Наиболее сильное перекрытие экологических ниш G. ribis и S.ribis наблюдалось во влажные годы (коэффициент Жаккара - 1,0). В связи с неблагоприятными условиями для реализации тактики выживания микромицетов, в засушливые года перекрытие экологических ниш G. ribis и S.ribis было на уровне 50 %.
Данные наблюдений за возбудителями септориоза и антракноза смородины черной представлены в таблице 2.
Таблица 2
Роль сорта в реализации экологической ниши S. ribis и G. ribis на смородине черной, среднее за 2006-2013гг.
|
Сорт |
Размер экологической ниши S. ribis, % |
Размер экологической ниши G. ribis, % |
||
|
лимиты |
среднее |
лимиты |
среднее |
|
|
Плотнокистная |
0,8÷14 |
7,6 |
0,2÷1,0 |
0,2 |
|
Черный жемчуг |
2,5÷25 |
12,5 |
0,6÷1,6 |
1,0 |
|
Любава |
12,5÷50 |
22,5 |
2,8÷6,2 |
4,1 |
|
Памяти Потапенко |
14÷75 |
45,8 |
1,0÷8,2 |
6,2 |
|
НСР05 |
|
0,39 |
|
0,34 |
Данные таблицы свидетельствуют, что на высокоустойчивом сорте Плотнокистная размеры экологических ниш S. ribis и G. ribis были в 6 и 31 раз меньше по сравнению с восприимчивым сортом Памяти Потапенко соответственно, и в 2,9 и 20,5 раз по сравнению со слабоустойчивым сортом Любава. Была выявлена достоверная корреляционная связь между развитием септориоза и антракноза по сортам (r = 0,95) и годам (r = 0,89). Размер экологической ниши S. ribis в засушливых условиях при снижении конкуренции и наличии большого числа свободных топических ниш был в 14 раз больше, чем у G. ribis, тогда как во влажные годы, характеризующиеся благоприятными для расселения и размножения обоих видов условиями, конкуренция существенно обострялась, и соотношение размеров реализованных ниш сдвигалось в пользу G. ribis в 1,6 раза.
На восприимчивых и слабоустойчивых сортах наблюдалось повреждение листьев и почек фитофагами, входящими в сообщество деструктивной биоты агроценозов смородины черной. При повреждении листьев тлей увеличивалась степень реализации экологической ниши G. ribis. Коэффициент корреляции размера экологической ниши G. ribis и Cr. ribis составил для восприимчивых сортов 0,64. Между G. ribis и Cr. ribis формировались взаимоотношения по типу протокооперации. Cr. ribis не оказывала воздействие на реализацию тактик Р (размножение), В (выживание), Т (трофические связи) S. ribis, проявляя нейтральные отношения (r = 0,18) с этим микромицетом. Клещ смородинный почковый (Cecidophyes ribis) повреждал почки смородины. Между фитопатогенами и C. ribis имели место нейтральные взаимоотношения из-за несовпадения экологических ниш. Антагонистические микроорганизмы, в частности ризосферные штаммы Ps. fluorescens, взаимодействовали с фитопатогенами по типу антагонизма.
При нанесении суспензии антагонистов на опавшие листья наблюдалось ограничение выживания S.ribis, повреждение пикнид и пикноспор, шло активное разложение растительных остатков. В результате уменьшалось число заражений листьев смородины черной S.ribis, снижалась скорость расширения экологической ниши микромицета (табл. 3).
Таблица 3
Влияние антагонистов при однократной обработке на паразитическую активность S.ribis на сортах смородины черной, среднее за 2008-2009, 2013 гг.
|
Группы сортов |
Вариант |
Размер экологической ниши, % |
Число некротических пятен / 1см2 |
||
|
lim |
среднее |
lim |
среднее |
||
|
Восприимчивые |
контроль |
46÷58 |
52,0 |
3,2÷6,2 |
5,4 |
|
антагонисты |
22÷34 |
28,2 |
2,2÷3,4 |
2,9 |
|
|
Слабоустойчивые |
контроль |
22÷42 |
36,0 |
2,4÷3,8 |
3,4 |
|
антагонисты |
8÷20 |
12,0 |
0,8÷1,8 |
1,0 |
|
|
Среднеустойчивые |
контроль |
14÷20 |
17,0 |
1,6÷2,6 |
2,2 |
|
антагонисты |
4÷12 |
8,0 |
0,6÷1,6 |
1,3 |
|
|
Высокоустойчивые |
контроль |
6÷12 |
8,1 |
0,7÷1,2 |
0,8 |
|
антагонисты |
1,2÷4 |
2,0 |
0,2÷0,5 |
0,3 |
|
|
НСР05 |
|
|
2,54 |
|
0,3 |
|
Степень влияния экологических факторов, % (по Снедекору): |
|||||
|
Сорт |
|
57,97 |
|
55,8 |
|
|
Антагонисты |
|
30,51 |
|
30,9 |
|
При нанесении суспензии антагонистов на листовой опад смородины черной число заражений или некротических пятен на вегетирующих листьях было в 2,4 раза ниже в среднем по группам сортов по сравнению с контролем. Особенно сильное подавление споруляции S.ribis и снижение числа заражений (в 3,4 раза) было отмечено в группе слабоустойчивых сортов. Однако дисперсионный анализ показал, что влияние антагонистов на число заражений оказалось в 1,8 раза ниже, по сравнению с влиянием сорта. На восприимчивых сортах антагонистам не удалось нормализовать ситуацию и размер экологической ниши S.ribis в конце вегетации был выше эпидемического порога, что отрицательно сказалось на продуктивности растений.
Для усиления воздействия Ps. fluorescens на ограничение паразитической активности S.ribis было проведено повторное нанесение суспензии антагонистов на вегетирующие листья при первых признаках паразитической активности микромицета. Выделяемые антагонистами биологически активные вещества обеспечивают снижение интенсивности размножения S. ribis на листьях разных по устойчивости сортов смородины черной (табл. 4).
Таблица 4
Влияние антагонистов при двойной обработке на интенсивность размножения S. ribis на сортах черной смородины, среднее за 2008-2009, 2013 гг.
|
Группы сортов |
Вариант |
Число пикнид / 1 некротическое пятно |
Число пикноспор / 1 пикниду |
Размер экологичес-кой ниши, % |
||
|
lim |
среднее |
lim |
среднее |
|||
|
Восприимчивые |
контроль |
11÷18 |
12,0 |
142÷324 |
228 |
52,0 |
|
антагонисты |
6÷12 |
8,0 |
94÷242 |
138 |
28,0 |
|
|
Слабо-устойчивые |
контроль |
6÷16 |
7,0 |
74÷312 |
195 |
36,0 |
|
антагонисты |
2÷4 |
2,2 |
38÷146 |
104 |
10,0 |
|
|
Средне-устойчивые |
контроль |
4÷16 |
6,5 |
48÷210 |
136 |
17,0 |
|
антагонисты |
0÷6 |
2,8 |
18÷94 |
76 |
8,0 |
|
|
Высоко-устойчивые |
контроль |
2÷8 |
3,5 |
34÷102 |
58 |
8,1 |
|
антагонисты |
0÷4 |
1,5 |
10÷72 |
38 |
2,1 |
|
|
НСР05 |
|
|
1,65 |
|
3,05 |
2,58 |
|
Степень влияния экологических факторов, % (по Снедекору): |
||||||
|
Сорт |
|
|
57,6 |
|
55,87 |
55,34 |
|
Обработка |
|
|
37,0 |
|
34,95 |
31,90 |
На группах слабоустойчивых и восприимчивых сортов размер экологической ниши в контроле соответствовал уровню умеренной эпифитотии. В группе восприимчивых сортов двойное воздействие антагонистов на популяцию S. ribis в период функционирования на опавших и вегетирующих листьях ограничило размер экологической ниши фитопатогена в 1,9 раза, однако не обеспечило радикального оздоровления растений. При нанесении суспензии Ps. fluorescens на листовой опад и вегетирующие листья число пикнид на пятно и число пикноспор на пикниду уменьшалось в 2 и 1,7 раз соответственно в среднем по группам сортов по сравнению с контролем. Сильнее всего подавление интенсивности размножения S.ribis антагонистами было выявлено в группе слабовосприимчивых сортов, где указанные параметры были в 3,2 и 1,9 раза ниже контрольных соответственно.
Выводы
- В агроценозах смородины черной было выявлено сообщество из 12 видов и групп организмов, среди которых наиболее распространенными в годы исследований были Septoria ribis, Gloeosporium ribis, Cecidophyes ribis, Cryptomyzus ribis, сапротрофные микроорганизмы филлопланы и ризосферы.
- В сообществе были выявлены следующие формы межпопуляционных взаимоотношений: паразитизм между смородиной и фитопатогенами (S. ribis, G. ribis), смородиной и фитофагами (C. ribis, Cr. ribis), конкуренция между фитопатогенами, антагонизм между фитопатогенными микромицетами и микроорганизмами Ps. fluorescens и Streptomyces spp., нейтрализм между смородинным почковым клещом (C. ribis) и фитопатогенными микромицетами, антагонистами и фитофагами; протокооперация проявилась между листовой галловой тлей (Cr. ribis) и эпифитной микрофлорой, неспецифическое угнетение между смородиной черной и эпифитными организмами.
- Конкурентные отношения G. ribis и S. ribis привели к ограничению размера экологической ниши G. ribis в 38 раз на устойчивом сорте Плотнокистная и в 7,4 раза - на восприимчивом сорте Памяти Потапенко по сравнению с S. ribis, что связано с более поздним инфицированием листьев смородины G. ribis. Была выявлена достоверная корреляционная связь между развитием септориоза и антракноза по сортам (r = 0,95) и годам (r = 0,89). Установлена более высокая конкурентная способность S. ribis по сравнению с G. ribis.
- Нанесение суспензии штаммов Ps. fluorescens на листовой опад ограничивало выживание S. ribis, вызвало повреждение пикнид и пикноспор, ускорило разложение растительных остатков, уменьшило размер экологической ниши фитопагена, особенно по группам слабо- и среднеустойчивых сортов.
- Двойное нанесение суспензии Ps. fluorescens на опавшие и вегетирующие листья ограничило интенсивность размножения S.ribis и размер его экологической ниши, особенно по группам слабо- и среднеустойчивых сортов.
Рецензенты:
Воробьева И.Г., д.б.н., доцент, заместитель директора по научной работе, ФГБОУ ВПО «РЭУ им. Г.В. Плеханова», Новосибирский филиал, Министерство образования и науки Российской Федерации, г. Новосибирск;
Соловьева А.Е., д.с.-х.н., консультант по научным исследованиям и разработкам в области естественных наук, ООО «Северный стиль», г. Новосибирск.
Библиографическая ссылка
Рябова А.А., Торопова Е.Ю. МЕЖПОПУЛЯЦИОННЫЙ АНАЛИЗ ФИТОЦЕНОЗОВ СМОРОДИНЫ ЧЕРНОЙ В СЕВЕРНОЙ ЛЕСОСТЕПИ ПРИОБЬЯ // Современные проблемы науки и образования. – 2014. – № 6. ;URL: https://science-education.ru/ru/article/view?id=16207 (дата обращения: 19.04.2024).