



Введение
Алтае-Саянская горная страна – наиболее высокая горная система Сибири с очень сложной структурой рельефа и значительным разнообразием ландшафтов [1, 5, 19]. Алтай и Саяны с давних пор освоены человеком, и к настоящему времени значительная часть их территории претерпела существенные антропогенные изменения в соответствии с преобладающими видами хозяйственной деятельности [18]. Сформированные таким образом местообитания чрезвычайно разнообразны, как по своей структуре, так и по степени нарушенности [2-4, 7, 8, 10-12, 15, 21, 23, 24].
Птицы – очень удобная модельная группа для географического мониторинга. Изменчивость орнитокомплексов относительно давно используют для оценки последствий антропогенной трансформаций ландшафтов, при этом могут быть использованы различные методологические подходы [17, 27, 28]. Комплексного анализа населения птиц антропогенных местообитаний региона ранее не проводили.
Цель исследования – выявить основные факторы, формирующие орнитокомплексы рассматриваемых местообитаний, многообразие антропогенных воздействий в пределах региона, а также детально изучить особенности пространственной структуры сообществ птиц.
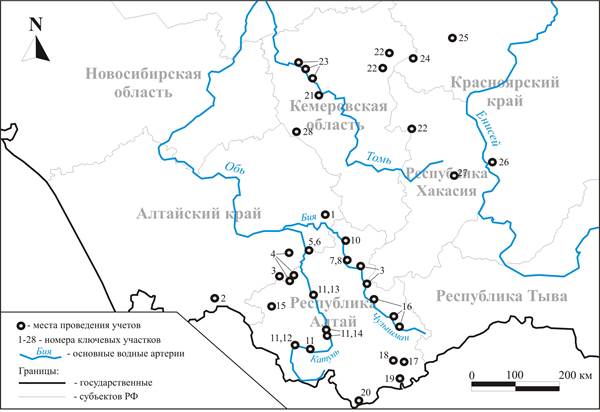
Рис. 1. Места проведения учетов птиц на территории Алтае-Саянской горной страны
Материалы и методы исследования
Анализ пространственной неоднородности населения птиц антропогенных ландшафтов проведен на основе материалов количественных учетов, накопленных в банке данных коллективного пользования ИСиЭЖ СО РАН. В итоговую выборку вошли как собственные материалы автора (рис. 1, ключевые участки 6, 10, 15), так и литературные данные, собранные другими исследователями [2, 3, 7, 10–12, 21, 23, 24]. Чтобы максимально снизить влияние миграционной активности на результаты расчетов, для анализа были использованы только данные учетов первой половины лета (с середины мая по середину июля). Маршрутные учеты проводили без ограничения ширины трансекта с последующим раздельным пересчетом полученных данных на площадь по средним групповым дальностям обнаружения интервальным методом [14, 17]. Общая протяженность маршрутов для 105 двухмесячных вариантов населения составила около 1500 км. Для выявления пространственно-типологической структуры и организации населения птиц использованы методы кластерного анализа. Варианты населения были разделены по коэффициентам сходства на классы таким образом, чтобы дисперсия матрицы коэффициента сходства, учитываемая этим разделением, была максимальна [22]. В качестве меры сходства использован коэффициент П. Жаккара [26] в модификации Р.Л. Наумова (1964). При построении графа сходства по полученному разбиению использован метод корреляционных плеяд [20]. В матрице межклассовых коэффициентов сходства выбирали максимальные значения связи. Полученные таким образом графы наилучшим образом иллюстрировали основные изменения населения и коррелирующие с ними факторы среды. Проверка правильности ориентации графа в факторном пространстве проведена методом многомерного (неметрического) шкалирования в программе STATISTICA 8.0 [30]. Оценка силы связи между факторами среды и пространственной изменчивостью населения птиц проведена с помощью метода качественной линейной аппроксимации матриц сходства [9]. Зависимость общей матрицы сходства населения птиц от матриц сходства каждого фактора среды оценивали на основании набора отдельных градаций факторов среды, выявленных при классификационных и структурных построениях. Сперва был проведен однофакторный анализ, далее рассчитана множественная регрессия по всем выявленным факторам и их сочетаниям (природно-антропогенным режимам). В качестве критерия вариабельности значений коэффициентов сходства принята дисперсия – средний квадрат отклонения от среднего по всей матрице значения [16]. Видовые названия птиц, за исключением маскированной трясогузки и седоголового щегла, приведены по А.И. Иванову [6]. Материалы обработаны с помощью программного обеспечения банка данных лаборатории зоологического мониторинга Института систематики и экологии животных СО РАН.
Результаты и обсуждение
Полученные результаты представлены в виде классификации населения птиц; структурного графа, отражающего основные тенденции пространственной неоднородности орнитокомплексов на уровне подтипов (рис. 2); оценки силы связи факторов среды с территориальной изменчивостью орнитокомплексов. Основное назначение классификации заключено в отражении сходства сообществ в виде иерархической схемы. В предлагаемой классификационной схеме для каждого таксона приведены лидирующие виды (первые пять по убыванию среднего обилия), их доля в населении (%), плотность населения (особей/км²), биомасса (кг/км²), видовое и фоновое богатство, а также фаунистический состав орнитокомплексов – доля представителей различных типов фауны в процентах [25].
Классификация населения птиц антропогенных местообитаний Алтае-Саянской горной страны
1 – застроенных местообитаний: домовый воробей и полевой воробьи (29 и 21), сизый голубь (18), большая синица (4), скворец (4); 1433/127; 167/49; транспалеарктов – 58, европейского и средиземноморского типов фауны – по 18;
1.1 – городов: сизый голубь (29), домовый и полевой воробьи (28 и 19), большая синица (5), горихвостка-лысушка (4); 2012/238; 68/31; транспалеарктов – 52, средиземноморского типа фауны – 29, европейского – 16;
1.1.1 – многоэтажной застройки крупных городов: домовый воробей (38), сизый голубь (30), полевой воробей (13), городская ласточка (3), горихвостка-лысушка (3); 1939/228; 43/23; транспалеарктов – 57, средиземноморского типа фауны – 30;
1.1.2 – многоэтажной застройки малых городов: сизый голубь (27), полевой воробей (19), большая синица (14), домовый воробей (14), маскированная трясогузка (6); 1667/182; 34/26; транспалеарктов – 40, средиземноморского типа фауны – 27, европейского – 25;
1.1.3 – двух и трехэтажной застройки: сизый голубь (66), домовый и полевой воробьи (22 и 7), скворец и большая синица (по 1); 4709/974; 17/13; средиземноморского типа фауны – 66, транспалеарктов – 30;
1.1.4 – одноэтажной застройки: полевой воробей (37), домовый воробей (22), большая синица (9), горихвостка-лысушка (9), галка (3); 1602/93; 53/34; транспалеарктов – 65, европейского типа фауны – 30;
1.2 – поселков и застроенных садов: домовый воробей и полевой воробьи (33 и 23), сизый голубь (11), скворец (6), маскированная трясогузка (4); 1303/93; 160/53; транспалеарктов – 64, европейского типа фауны – 16, средиземноморского – 11;
1.2.1 – поселков лесных крупных: домовый и полевой воробьи (32 и 23), сизый голубь (12), скворец (5), горихвостка-лысушка (4); 1033/73; 92/43; транспалеарктов – 61, европейского типа фауны – 19, средиземноморского – 12;
1.2.2 – степных крупных: домовый воробей (41), сизый голубь (31), полевой воробей (7), маскированная трясогузка (6), скворец (3); 987/118; 65/31; транспалеарктов – 51, средиземноморского типа фауны – 32;
1.2.3 – лесных средних: домовый воробей и полевой воробей (21 и 16), деревенская ласточка (9), скворец (8), маскированная трясогузка (6); 1017/51; 104/50; транспалеарктов – 58, европейского типа фауны – 25;
1.2.4 – лесостепных: домовый и полевой воробьи (71 и 14), горихвостка-лысушка (6), серая ворона (2), садовая камышевка (1); 2393/103; 32/19; транспалеарктов – 86, европейского типа фауны – 11;
1.2.5 – лесных малых: домовый воробей (20), сизый голубь (17), деревенская ласточка (6), галка (5), чечевица (5); 1046/93; 75/48; транспалеарктов – 49, европейского типа фауны – 19, средиземноморского – 17;
1.2.6 – степных малых: домовый и полевой воробьи (37 и 28), сизый голубь (13), скворец (5), маскированная трясогузка (3); 1766/137; 106/43; транспалеарктов – 70, средиземноморского типа фауны – 13, европейского – 11;
1.2.7 – садов застроенных: полевой воробей (49), скворец (20), домовый воробей (8), варакушка (6), коноплянка (2); 1613/67; 50/32; транспалеарктов – 64, европейского типа фауны – 30;
1.3 – промзон, карьеров, отвалов: полевой и домовый воробьи (29 и 17), сизый голубь (6), маскированная трясогузка (5), варакушка (4); 370/23; 77/36; транспалеарктов – 67, европейского типа фауны – 14;
1.4 – городских парков и скверов: большая синица (26), горихвостка-лысушка (15), полевой воробей (10), зяблик (7), сизый голубь (5); 1353/105; 40/34; европейского типа фауны – 71, транспалеарктов – 17;
2 – кордонов и зимних пастушьих стоянок: маскированная трясогузка (18), большая синица (9), домовый воробей (7), большая синица (7), полевой воробей (6); 223/11; 55/33; европейского типа фауны – 43, транспалеарктов – 17;
2.1 – кордонов: маскированная трясогузка (19), горихвостка-лысушка (10), домовый воробей (7), большая синица (7), полевой воробей (5); 274/13; 41/32; европейского типа фауны – 46, транспалеарктов – 17;
2.2 – стоянок: полевой и снежный воробьи (22 и 13), клушица (13), каменный воробей (10), сизый голубь (9); 69/6; 22/10; монгольского типа фауны – 26, транспалеарктов - 23, средиземноморского типа фауны – 19, тибетского – 14;
3 – полуоблесенных местообитаний: полевой воробей (12), бормотушка (10), черноголовый чекан (9), полевой жаворонок (6), сорока (6); 539/47; 159/52; транспалеарктов – 34, европейского типа фауны – 33, китайского – 11, средиземноморского – 10;
3.1 – сосновых посадок и ивовых зарослей гидроотвалов: сорока (11), рябинник (8), полевой воробей и черноголовый чекан (по 8), бормотушка (5); 547/45; 87/47; европейского типа фауны – 44, транспалеарктов – 25, сибирского типа фауны – 17;
3.2 – лесополос: бормотушка и полевой воробей (по 17), полевой жаворонок (11), грач и дубровник (по 7); 734/75; 74/39; транспалеарктов – 42, европейского типа фауны – 23, средиземноморского – 17, китайского – 12;
3.2.1 – широких: полевой жаворонок (26), бормотушка (25), черноголовый чекан (10), желтая трясогузка (7), дубровник (4); 797/50; 57/38; транспалеарктов – 48, средиземноморского типа фауны – 25;
3.2.2 – узких: полевой воробей (25), бормотушка (13), грач (11), дубровник (9), сорока (7); 702/88; 74/39; транспалеарктов – 38, европейского типа фауны – 31, китайского – 13, средиземноморского – 13;
3.3 – полей, лугов, выпасов в сочетании с перелесками, садов незастроенных: черноголовый чекан (15), лесной конек (10), чечевица (7), серая славка и обыкновенная овсянка (по 5); 360/22; 138/53; европейского типа фауны – 43, транспалеарктов – 28, китайского типа фауны – 12;
3.3.1 – полей, лугов, выпасов в сочетании с перелесками: черноголовый чекан (15), лесной конек (10), чечевица (7), серая славка (6), полевой воробей (5); 352/22; 130/48; европейского типа фауны – 43, транспалеарктов – 29, китайского типа фауны – 12;
3.3.2 – садов незастроенных: черноголовый чекан (15), лесной конек (10), чечевица (9), жулан (7), полевой конек (6); 434/25; 58/37; европейского типа фауны – 45, транспалеарктов – 22, китайского типа фауны – 14;
4 – открытых местообитаний: полевой жаворонок (18), каменка-плясунья (14), садовая овсянка и, степной конек (по 7), черноголовый чекан (5); 283/17; 143/38; транспалеарктов – 35, монгольского типа фауны – 26, европейского – 23;
4.1 - полей степных горнодолинных и лугов, покосов: полевой жаворонок (39), садовая овсянка (10), черноголовый чекан и лесной конек (по 6), береговая ласточка (4); 197/12; 77/26; транспалеарктов – 57, европейского типа фауны – 23;
4.1.1 – полей степных горнодолинных: полевой жаворонок (36), садовая овсянка (15), черноголовый чекан (7), береговая ласточка (5), каменка-плясунья (4); 185/10; 62/22; транспалеарктов – 52, европейского типа фауны – 26, монгольского – 10;
4.1.2 – лугов, покосов: полевой жаворонок (47), лесной конек (11), травник (6), перепел (6), черноголовый чекан (4); 234/20; 34/21; транспалеарктов – 67, европейского типа фауны -16;
4.2 - выпасов увлажненных: степной конек (27), черноголовый чекан (8), полевой жаворонок (7), каменка-плясунья (6), седоголовый щегол (5); 429/38; 82/34; транспалеарктов и монгольского типа фауны – по 35, европейского – 18;
4.3 – выпасов сухих, остепненных: каменка-плясунья (30), каменка (6), садовая овсянка (5), желтоголовая трясогузка и рогатый жаворонок (по 4); 381/16; 89/40; монгольского типа фауны – 39, европейского – 25, транспалеарктов – 13.
Представленной классификацией аппроксимируется 47 % дисперсии коэффициентов сходства между рассматриваемыми сообществами птиц (коэффициент множественной корреляции 0,7). Население антропогенных местообитаний на наиболее высоком уровне классификации (тип населения) проявляет сопряженность с самыми общими ландшафтными характеристиками – наличием застройки и степенью облесенности территории. Однако разбиение типов на подтипы в каждом случае обусловлено специфичным набором факторов. Для населения птиц застроенных территорий (тип 1) характерно наибольшее внутреннее разнообразие, что отражено в максимальном числе таксонов низкого ранга (4 подтипа и 13 классов). Подразделенность на подтипы соответствует характеру и степени застроенности и в меньшей степени особенностям природного окружения. Разбиение населения птиц городов (подтип 1.1) на уровне классов не проявляет сопряженности с ландшафтным окружением или провинциальными особенностями местообитаний, отражая лишь размеры города и этажность застройки. В то же время население поселков (подтип 1.2) на уровне классов демонстрирует сопряженность как с размерами населенных пунктов, так и с характером их природного окружения. Выделение подтипов в населении птиц лесных кордонов и пастушьих стоянок (тип 2) также связано с особенностями окружающих природных ландшафтов. Разбиение населения птиц полуоблесенных местообитаний (тип 3) на подтипы совпадает с характером и степенью мозаичности местообитаний, поскольку для лесополос (подтип 3.2) и посадок на гидроотвалах (подтип 3.1) мозаичность определяет их внешнее окружение, тогда как мозаичность полей, лугов и выпасов в сочетании с перелесками (подтип 3.3) имеет внутренний характер [29]. Варианты сообществ, составляющих тип 4 (орнитокомплексы открытых местообитаний), объединены в три подтипа в соответствии со степенью увлажнения и характером антропогенной нагрузки (выпас, распашка, сенокошение).
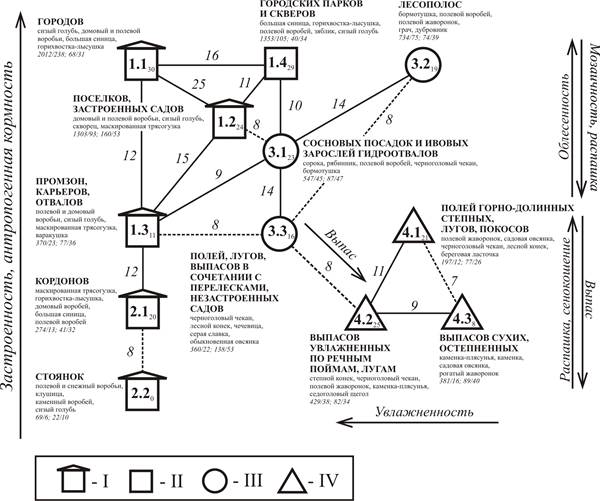
Рис. 2. Пространственно-типологическая структура населения птиц антропогенных местообитаний Алтае-Саянской горной страны в первой половине лета на уровне подтипа населения.
Подтипы сообществ птиц местообитаний: I – застроенных, II – облесенных, III – полуоблесенных, IV – открытых; цифры внутри фигур – номера типов и подтипов по классификации орнитокомплексов, нижний индекс – внутриклассовое сходство; величина межклассового сходства приведена около линии, соединяющей классы; сплошная линия – межклассовые связи при пороге значимости 9 единиц, прерывистая – связи ниже порога значимости; в характеристике классов указаны: пять лидирующих видов (первых по обилию); плотность населения (особей/км²), биомасса (кг/км²) и число видов (встреченных/фоновых); стрелки – направления градиентов основных структурообразующих трендов среды (в сторону усиления влияния).
Пространственно-типологическая структура наглядно демонстрирует основные направления территориальной изменчивости населения птиц и определяющие ее факторы среды (рис. 2). Анализ пространственно-типологической структуры населения птиц антропогенных местообитаний Алтае-Саянской горной страны позволяет выделить основные тренды изменчивости населения и соответствующие им факторы среды: застроенность, антропогенная кормность, облесенность, мозаичность, увлажненность, распашка, выпас, сенокошение.
В группе застроенных местообитаний (подтипы населения 1.1-1.3 и 2.1-2.2) в вертикальном направлении четко выражен тренд, отражающий степень застроенности, размер застроенной территории и сопряженную с этим антропогенную кормность местообитаний. Его проявления также прослежены в последовательном возрастании показателей суммарного обилия и биомассы в ряду от слабо застроенных местообитаний к полностью урбанизированным. Поскольку в застроенных местообитаниях лидируют чаще всего синантропы, изменчивость состава лидеров по обилию касается, прежде всего, порядка их лидерства и в меньшей степени состава других видов, распространение которых менее связано с человеком.
Изменчивость по облесенности отражена на графе по вертикали от подтипа 3.3 к подтипу 3.1, а затем к 1.4 и 3.2 и сопряжена с относительной площадью древостоев, как естественного, так и искусственного происхождения. На увеличение облесенности птицы также реагируют увеличением суммарного обилия и биомассы, которые постепенно возрастают от полей, лугов, выпасов в сочетании с перелесками и садов до городских парков и скверов. Помимо облесенности, в ряду рассматриваемых местообитаний заметно влияние мозаичности местообитаний, направленное в значительной мере в противоположную сторону по отношению к облесенности. В отличие от застроенных территорий, где состав преобладающих по обилию видов относительно стабилен и однообразен, облесенные и полуоблесенные территории гораздо разнообразнее по лидерам. Различия в структуре древесно-кустарникового покрова и степени мозаичности обуславливают принципиально разный состав преобладающих по обилию видов в облесенных и полуоблесенных местообитаниях.
Влияние выпаса, распашки и сенокошения отражено на графе в вертикальном ряду (от подтипов 4.2 и 4.3 к 4.1) и проявляется, прежде всего, в более низких показателях обилия и биомассы по сравнению с нераспаханными местообитаниями, где выпасают скот. Кроме того, перечисленные таксоны формируют горизонтальный ряд, совпадающий с изменчивостью местообитаний по степени их увлажненности. Для населения наиболее увлажненных участков (подтип 4.2) свойственны максимальные показатели обилия и биомассы в пределах типа, поскольку в них относительно высока кормообеспеченность, значительное обилие видов-посетителей, а в числе фоновых видов присутствуют птицы средних и крупных размеров. Вместе с тем, максимальные значения видового и фонового богатства характерны для относительно сухих местообитаний (подтип 4.3), что связано с разнообразием форм рельефа и растительности этих участков, а также с влиянием экспозиционных различий. Необходимо отметить, что сходство орнитокомплексов, отнесенных к подтипам 4.1-4.3 со всеми остальными сообществами ниже порогового значения, в связи с меньшим обилием синантропов и своеобразием видового состава их населения. Список лидеров определяют описанные выше факторы и некоторыми провинциальные особенности.
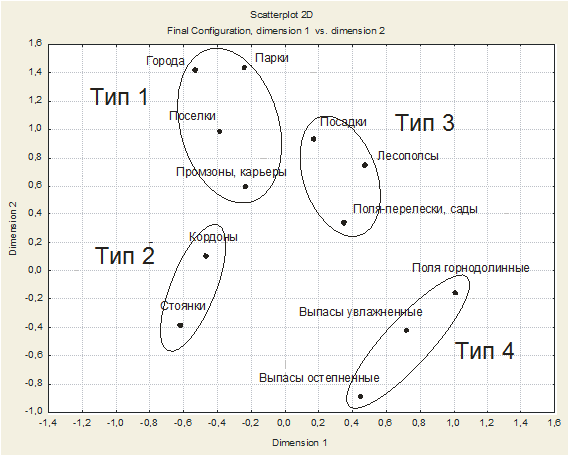
Рис. 3. Результат многомерного (неметрического) шкалирования по подтипам населения птиц, двухмерная проекция.
Многомерное (неметрическое) шкалирование подтверждает правильность ориентации графа в факторном пространстве (рис. 3), группировку подтипов населения в более высокие иерархические категории, а также значительную обособленность типов 2 и 4 от соседних таксонов.
Для оценки силы связи неоднородности населения птиц региона с факторами среды заданы 14 факторов, по которым прослежена коррелятивная связь с изменчивостью населения птиц. В таблице 1 приведены те проанализированные факторы, значимость связи которых составила не менее 1 % учтенной дисперсии.
Наиболее значимая группа факторов отражает различные аспекты застроенности территории. В частности, максимальный показатель учтенной дисперсии – 41 %, учитывает фактор, определяющий характер застроенности (сплошной или точечной). При этом с фактором застроенности как таковой, а также ее этажности связана меньшая доля дисперсии (по 33 %).
Высокое значение учтенной дисперсии по характеру застроенности косвенно свидетельствует о важности таких параметров, как площадь застроенной территории и удаленности ее центра от границ с окружающими незастроенными местообитаниями. Очевидно, что в точечных вариантах застройки, таких как кордоны и стоянки, удаленность центра от границ гораздо меньше, чем в городах и поселках, и формирование населения птиц таких местообитаний в значительно большей мере определяют условия природного окружения, нежели характеристики самих участков.
Таблица 1
Оценка силы связи факторов среды и неоднородности населения птиц антропогенных местообитаний Алтае-Саянской горной страны
|
Фактор |
Учтенная дисперсия, % |
|
Характер застроенности (сплошная/точечная) |
41 |
|
Размер населенного пункта |
38 |
|
Застроенность |
33 |
|
Этажность застройки |
33 |
|
Природное окружение |
32 |
|
Провинциальность |
12 |
|
Облесенность |
7 |
|
Все факторы |
58 |
|
Режимы по классификации |
47 |
|
Режимы по структуре |
49 |
|
Режимы по классификации и структуре |
51 |
|
Все факторы и режимы, вместе взятые |
60 |
Размер населенного пункта объясняет 38 % учтенной дисперсии. Этот фактор скоррелирован с общим характером застройки, поскольку взаимосвязан с площадью местообитания и удаленностью центра от границ с окружающими ландшафтами.
Различия в окружении населенных пунктов снимают 32 % дисперсии. Столь высокая доля отражает участие «вобранных» видов в формировании населения соответствующих местообитаний.
Далее по значимости следует фактор провинциальности – 12 % учтенной дисперсии; в нем выделено наибольшее число градаций (11). Сходный показатель (10 %) по провинциальности отмечен для всех ландшафтов Алтайской горной страны [24]. Несмотря на значительно большее внутреннее сходство антропогенных местообитаний по сравнению с природными и слабо нарушенными, столь близкие значения учтенной дисперсии по двум различным выборкам свидетельствуют о почти идентичном вкладе провинциальных особенностей территорий в формирование пространственной неоднородности населения птиц антропогенных и слабонарушенных ландшафтов в указанных регионах.
Облесенность, заданная тремя градациями (высокая, средняя и низкая), снимает 7 % учтенной дисперсии. Следует отметить, что в анализируемой выборке отсутствуют настоящие лесные местообитания и облесенность в большинстве случаев представлена естественным окружением соответствующих местообитаний, либо древесно-кустарниковыми массивами искусственного происхождения внутри них (парки, скверы, лесополосы).
Выявленная система факторов среды, определяющая пространственное распределение птиц региона, объясняет 58 % исходной дисперсии матрицы коэффициентов сходства, а всеми факторами и режимами можно объяснить 60 % дисперсии коэффициентов сходства (коэффициент множественной корреляции 0,77).
Таким образом, набор выявленных факторов, в различной мере определяющих пространственную неоднородность населения птиц антропогенных местообитаний, достаточно велик. Однако лишь небольшая их часть объясняет высокую долю учтенной дисперсии. Наиболее значимыми факторами оказались те, которые отражают степень и характер застроенности, размер населенного пункта, а также особенности природного окружения. Остальные факторы, несмотря на заметные проявления на локальном уровне, не вносят весомого вклада в общую картину пространственной неоднородности населения.
Заключение
По результатам анализа классификации населения и его пространственно-типологической структуры выявлен набор факторов среды, определяющих основные направления изменчивости сообществ птиц. Структура населения антропогенных местообитаний Алтае-Саянской горной страны формируется, прежде всего, в соответствии с комплексным проявлением застроенности и сопряженной с ней антропогенной кормности территории. Вместе с тем, влияние особенностей природного окружения так же велико и по численным значениям учтенной дисперсии ненамного уступает проявлениям застроенности. В то же время, несмотря на значительное разнообразие природных ландшафтов региона, провинциальная специфика объясняет лишь 12 % учтенной дисперсии, что близко к аналогичному показателю для полной ландшафтной выборки по Алтайской части горной страны. Ряд факторов, весьма значимых на локальном уровне (распашка, выпас, мозаичность), не вносят существенного вклада в общую картину организации населения.
Исследования выполнены в рамках проекта РФФИ номер 13-04-00582.
Рецензенты:
Вартапетов Л.Г., д.б.н., зам. директора по науке, Институт систематики и экологии животных СО РАН, г. Новосибирск.
Литвинов Ю.Н., д.б.н., зам. директора по науке, Институт систематики и экологии животных СО РАН, г. Новосибирск.
Библиографическая ссылка
Хайдаров Д.Р., Богомолова И.Н. ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ ОРГАНИЗАЦИЯ НАСЕЛЕНИЯ ПТИЦ АНТРОПОГЕННЫХ МЕСТООБИТАНИЙ АЛТАЕ-САЯНСКОЙ ГОРНОЙ СТРАНЫ // Современные проблемы науки и образования. – 2013. – № 6. ;URL: https://science-education.ru/ru/article/view?id=11351 (дата обращения: 19.04.2024).