



Введение
Апоптоз представляет собой один из вариантов программируемой клеточной гибели. Индукция апоптоза и активация проапоптотических белков ведет к активации каспаз (цистеиновых протеаз) [1,6], которые делятся на инициаторные (2, 8, 9, 10) и эффекторные каспазы (3, 6, 7). Активация эффекторных каспаз является конечной стадией перед запуском протеолитического каскада реакций, ведущего к гибели клетки. Маркером более ранних событий апоптоза может служить детекция активации каспазы-8 при внешнем (рецепторном) пути активации апоптоза, которая происходит с поверхности клеточной мембраны при взаимодействии специфических рецепторов с растворимыми молекулами семейства фактора некроза опухоли (ФНО-подобными лигандами). Кроме того, недавно продемонстрирована роль каспазы 8 как инструмента некроза [2,4,7]. Таким образом, детектируя одновременно активацию каспазы-8 и каспазы-3, можно дискриминировать пути активации клеточной гибели: внешний или внутренний путь активации каспазы-3 или же активация другого пути программируемой клеточной гибели или клеточного ответа. Кроме того, активация каспаз не всегда ведет к гибели клетки. На некоторых моделях показано, что активация инициаторных каспаз связана не только с апоптозом, но и с противовоспалительным ответом клетки [3,5]. Поэтому регистрация ранних событий апоптоза, определяемых активацией каспазы-8 в отсутствие последующей активации каспазы-3, может рассматриваться как маркер запуска иных программируемых процессов в клетке.
Цель исследования
Целью данной работы стало изучение свойств генетически кодируемого FRET-сенсора каспазы-8 TagRFP-IETD-KFP, представляющего собой пару флуоресцентных белков TagRFP и KFP, соединенных гибким полипептидным линкером GTGGSGGIETDGTGGSGDPPVAT, содержащим тетрапептид IETD, специфически распознаваемый и гидролизуемый каспазой-8.
Материалы и методы
Очистка полученного сенсора
Клетки E.coli штамм BL21(DE3), содержащие белок TagRFP-IETD-KFP, осаждали центрифугированием (20 мин при 4 тыс. об/мин, 4ºС), суспендировали в буфере 20 мМ Трис´HCl, рН 7.8, 0.15М NaCl и разрушали с помощью френч-пресса. После разрушения клеток очистка полученного лизата проводилась с помощью гель-фильтрационной хроматографии на носителе Superdex 200 10/300 GL с использованием хроматографа AKTA purifier производства компании GE Healthcare (Швеция). Скорость элюирования составляла 0.5 мл/мин. Рабочий буфер: 150 мМ NaCl; 20 мМ Tris·HCl pH = 8,7. Регистрация поглощения выполнялась на двух длинах волн – 280 и 560 Целевой белок собирали по поглощению на 560 нм.
Калибровка колонки была выполнена с помощью стандартов для гель-фильтрации производства компании BioRad (Thyroglobulin (bovine) – 670 kDal γ-globulin (bovine) – 158 kDa; Ovalbumin (chicken) – 44 kDa; Myoglobin (horse) – 17 kDa; Vitamin B12 – 1,35 kDa).
Калибровочная кривая была получена путем построения зависимости коэффициента распределения Kav = (Ve-V0)/(Vt-V0) от десятичного логарифма молекулярной массы (Mr) стандартов, где Ve – объем элюирования, V0 – мертвый объем и Vt – общий объем колонки. Молекулярную массу белка определяли методом электрофореза в денатурирующих условиях по Лэммли в 12 % ПААГ на приборе MiniProtean II (Bio-Rad, США), с использованием набора маркеров молекулярных масс белков (Fermentas, США).
Измерение гидродинамического радиуса исследуемых сенсоров методом динамического светорассеивания
Для измерения гидродинамического радиуса использовали установку DynaPro Titan и программного обеспечения Dynamics 6.6.7.9 производства Wyatt Technology (США). Измерения проводили при температуре образца 25 °С, интенсивность лазерного облучения составляла 100 %. Для интерпретации результатов использовали предустановленную модель (MW-R model) «Globular Proteins». Для каждого измерения, в соответствии с рекомендациями производителя, было установлено время накопления сигнала 10 с и 10 измерений на один эксперимент. Определение олигомерного состояния проводили на основе данных о гидродинамическом радиусе исследуемого белка с помощью программного обеспечения Dynamics 6.6.7.9 (Wyatt Technology, США).
Определение эффективности переноса энергии в конструкции TagRFP-IETD-KFP
Спектры поглощения образцов TagRFP и конструкции TagRFP-IETD-KFP в буфере 150мМ NaCl; 20 мМ Трис-НСl рН 8,7 регистрировались на спектрофотометре Cary 300 (Varian,USA). Затем на спектрофлуориметре Cary Eclipse (Varian, USA) регистрировались спектры флуоресценции этих же образцов при возбуждении флуоресценции на длине волны 532 нм. На основании полученных спектров по формуле (1) была рассчитана эффективность переноса энергии в конструкции TagRFP-IETD-KFP.
E= 1 - n = 1- (Sem/ DTagRFP)/(S/D) (1),
Где Sem – площадь под спектром эмиссии флуоресценции конструкции, DTagRFP – значение оптической плотности TagRFP в конструкции на длине волны 532 нм, S – площадь под спектром эмиссии флуоресценции индивидуального TagRFP, D – значение оптической плотности индивидуального TagRFP на длине волны 532 нм.
Для нахождения DTagRFP было проведено разложение спектра поглощения TagRFP на два пика с помощью разложения Гаусса и определены положения и полуширины пиков. Спектры поглощения конструкции разложили с помощью четырёх Гауссов, после чего просуммировали оптические плотности каждой из двух компонент TagRFP на длине волны 532 нм в спектре поглощения конструкции.
Определение эффективности расщепления генетически кодируемого субстрата TagRFP-IETD-KFP
Белок слияния был отдиализован к буферу 100 mM NaCl, 50 mM HEPES, 10 mM DTT, 1 mM ЭДТА, 10 % глицерин, 0,5 % CHAPS, pH 7,4. Концентрация сенсора, используемая в эксперименте, равнялась 1,4 мкМ. К 93 мкл раствора субстрата добавляли 7 мкл каспазы-8 (100 ед/мкл), тщательно перемешивали и инкубировали в течение 24 часов при температуре 37 оС. Измерения спектров флуоресценции проводили на спектрофлуориметре Cary Eclipse (Varian, USA) при возбуждении светом с длиной волны 532 нм.
Для измерения времени жизни флуоресценции субстратов использовался флуоресцентный спектрометр с временным разрешением FluoTime 200 (Picoquant GmbH, Германия). Кинетики затухания флуоресценции белков обрабатывались по уравнению (1) с помощью программного обеспечения FluoFit (Picoquant Gmbh, Германия):
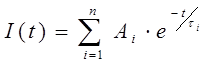 (1)
(1)
Расщепление исследуемого сенсора было также продемонстрировано с помощью электрофореза в денатурирующих условиях по Лэммли в 15 % ПААГ на приборе MiniProtean II (Bio-Rad, США), с использованием набора маркеров молекулярных масс белков (Fermentas, США).
Результаты и обсуждение
Характеристика степени созревания сенсора
По данным ПААГ-электрофореза в элюате находится достаточно чистый образец TagRFP-IETD-KFP. При этом наблюдаются две основные полосы – 37 и 18 кДа, в то время как в районе 26 кДа белковых фракций нет. Поскольку при созревании KFP в полипептидной цепи образуется разрыв, наблюдаемые полосы можно сопоставить соответственно форме субстрата с созревшим KFP (37 кДа) и фрагменту KFP, образовавшимся в результате разрыва полипептидной цепи (18 кДа).
Определение олигомерного состояния сенсора
Результаты гель-фильтрации говорят о том, что субстрат TagRFP-IETD-KFP в основном выходит в виде фракции, молекулярная масса которой составляет около 236 кДа, следовательно, так как масса цельной конструкции составляет 55 кДа, белок выходит в форме тетрамера.
В результате экспериментов по динамическому светорассеянию было вычислено, что гидродинамический радиус белка TagRFP-IETD-KFP равен 5,9 нм, что в приближении сферической модели соответствует молекулярной массе, равной 213 кДа, соответствующей тетрамерной форме белка. Таким образом, белок слияния TagRFP-IETD-KFP представляет собой тетрамер.
Определение эффективности переноса энергии
Для оценки эффективности переноса энергии в сенсоре TagRFP-IETD-KFP было найдено отношение квантовых выходов TagRFP в составе сенсора и в виде индивидуального белка: n = (Sem/DTagRFP532)/(S/D) = 44 %, т.е. в полученной конструкции происходит индуктивно-резонансный перенос энергии, эффективность которого составляет E = 100 % - 44 % = 56 %.
Гидролиз сенсора TagRFP-IETD-KFP каспазой-8 in vitro
В соответствии с рисунком 1 интенсивность флуоресценции возрастает приблизительно на 60 %.
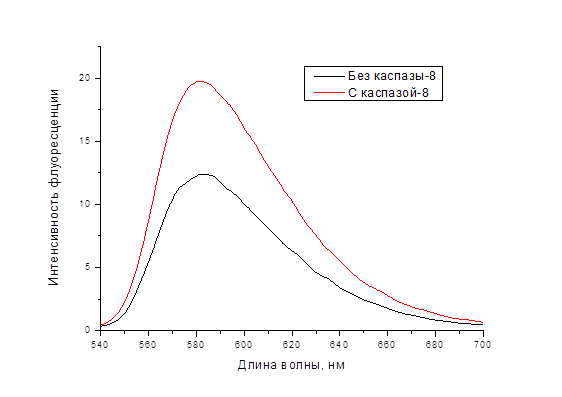
Рисунок 1. Спектр флуоресценции конструкции TagRFP-IETD-KFP до (чёрная линия) и после (красная) обработки каспазой-8
Таким образом, спектры флуоресценции свидетельствуют об эффективном расщеплении линкера под действием каспазы-8.
Детекцию гидролиза TagRFP-IETD-KFP каспазой-8 проводили также путем измерения времени жизни флуоресценции донора до и после добавления фермента. Кинетика затухания флуоресценции цельного субстрата описывается с помощью двух экспонент с временами жизни 2,4 и 1,3 нс, характеризующих расщепленный и цельный субстрат соответственно. Соотношение этих компонент до добавления каспазы-8 составляет 29 % к 71 %. В результате гидролиза каспазой-8 соотношение компонент изменяется до 50 % к 50 %, что также подтверждает расщепление субстрата и демонстрирует возможность детекции активности каспазы-8 по изменению времени жизни образца в соответствии с таблицей 1.
Таблица 1. Времена жизни и предэкспоненциальные множители для цельного и расщепленного субстрата TagRFP-IETD-KFP
|
|
τ1, нс |
τ2, нс |
A1 |
A2 |
|
Без каспазы |
2,2 |
1,1 |
1816 (29 %) |
4475 (71 %) |
|
С каспазой |
2,3 |
1,1 |
2833 (50 %) |
2865 (50 %) |
Таким образом, кинетика затухания флуоресценции конструкции TagRFP-IETD-KFP описывается двумя экспонентами, и в результате гидролиза каспазой-8 изменяется соотношение их амплитуд, что позволяет детектировать активность фермента. Детекцию активности каспазы-8 с помощью сенсора TagRFP-IETD-KFP можно проводить по изменению интенсивности флуоресценции и по времени жизни флуоресценции.
Работа выполнена при финансовой поддержке Министерства образования и науки по государственному контракту № 14.512.11.0023.
Рецензенты:
Королева О. В., д.б.н., зав. лабораторией Молекулярных основ биотрансформаций Института биохимии им. А. Н. Баха РАН, г.Москва.
Шишкин С. С., д.б.н., профессор, зав. лабораторией Биомедицинских исследований Института биохимии им. А. Н. Баха РАН, г.Москва.