



Природные популяции земноводных являются чувствительными индикаторами комплексного загрязнения окружающей среды, влияющей на репродуктивную систему, выживаемость, стабильность развития, генетическую структуру, ценотические связи и коэволюционные взаимоотношения [6]. Исследование функционально-физиологических перестроек в организме амфибий дает возможность получать информацию о механизмах поддержания устойчивости в условиях урбанизации, адаптивном потенциале популяций и возможности оценить перспективы их жизнедеятельности в конкретных биотопах Красноярского края. Целью настоящего исследования явилось изучение специфики морфоцитохимических показателей клеточных популяций VI слоя крыши среднего мозга озерной лягушки (Pelophylax ridibundus Pall.) в зависимости от уровня антропогенной трансформации местообитаний.
Материалы и методы
Исследования проведены на озерных лягушках (Pelophylax ridibundus Pall.), отловленных в июне 2008–2009 гг. в популяциях с территории Шарыповского района Красноярского края, который характеризуется значительным уровнем загрязнения [3]. В соответствии с типизацией В.Л. Вершинина [6], комплексно отражающей степень урбанизации, пруд п. Шарыпово относится к зоне малоэтажной застройки, а фоновая территория (где собрана контрольная выборка) выбрана в 23 км от п. Шарыпово, пойма р. Берешь Шарыповского района. Объем материала – 10 особей из зоны малоэтажной застройки (группа 1) и 7 – из загородной популяции (группа 2).
Забор образцов головного мозга животных осуществлялся в соответствии с приказом Министерства здравоохранения и социального развития РФ от 23.08.2010 № 708н г. Москва «Об утверждении правил лабораторной практики». Изъятый мозг фиксировали по стандартной гистологической методике для нервной ткани с последующим изготовлением серийных срезов на санном микротоме «Slide 2002». Готовые срезы, толщиной 5–7 мкм, окрашивали на выявление рибонуклеопротеидных комплексов тионином по Нисслю в модификации И.В. Викторова [2] и на содержание общего белка амидочерным (АЧ 10Б) [4]. Крыша среднего мозга земноводных, tectumopticum, состоит из поверхностного, центрального и перивентрикулярного слоев. Последние два слоя (центральный и перивентрикулярный) объединяются в так называемое центральное серое вещество. В перивентрикулярный слой входят 1–6 слои крыши среднего мозга [7]. В данной работе изучены эфферентные нейроны VI наружного зернистого слоя крыши среднего мозга земноводных, идентичные IV слою среднего мозга костных рыб. Средний мозг является высшим интегрирующим центром и обеспечивает регуляцию и координацию движений амфибий. Дифференцировку структур проводили по атласам мозга земноводных [10]. Изучены характеристики клеток – площадь сечения: тела – Sт, цитоплазмы – Sц, ядра – Sя и структурный ядерно-цитоплазматический коэффициент (сЯЦК), отношение Sя / Sц. Проанализированы плотность распределения нейронов и глиоцитов на единицу фиксированной площади (1 мм2). Подсчитано число нейронов с разной степенью хроматофилии цитоплазмы: нормохромные, гипохромные и гиперхромные [5]. На препаратах, окрашенных амидочерным 10Б, с помощью микроскопа Zeiss Axioskop со встроенной видеокамерой и прилагаемым программным обеспечением, в нейронах VI слоя крыши среднего мозга количественными методами определено суммарное содержание и концентрация общих водонерастворимых белков, составляющих основу сухого веса клеточных структур [8]. Полученные результаты обработаны непараметрическим критерием Манна – Уитни. Значимыми принимали значения при р < 0,05. Статистические данные представлены в виде Ме [25; 75].
Результаты исследования
Озерная лягушка на территории Красноярского края является инвазивным видом, появившимся в результате случайного заноса, вероятно, с молодью карпа из Европейской части страны [9]. Как полуводный вид P. ridibundus заселяет проточные и стоячие водоемы (от мелких луж, речек до крупных рек и водохранилищ). Она предпочитает открытые, хорошо прогреваемые водоемы с густой травяной растительностью [1]. P. ridibundus характеризуется высоким окрасочным и пигментным полиморфизмом. Она толерантна к повышенным концентрациям растворенных в воде солей (0,9–8,3 %) и способна обитать в водоемах, загрязненных бытовыми отходами и удобрениями, в окрестностях крупных металлургических и химических предприятий, в условиях, к которым аборигенные виды амфибий не способны адаптироваться [6].
Популяции эфферентных нейронов VI слоя крыши среднего мозга озерной лягушки группы 2 характеризуются наибольшей площадью профильного поля тела клетки в сравнении с лягушками1 группы. В кариохромных клетках данного слоя преобладают значения площади ядра в 1,7 раза у особей группы 1 и 1,5 раза у группы 2 над показателями площади цитоплазмы. Варьирование показателей Sя связано с реактивностью нейронов, обеспечивающих поддержание относительного постоянства внутренней среды организма, необходимой для выполнения конкретных функций, требующих оптимального пластического и энергетического баланса в клетках. В изученных биотопах в нейронах VI слоя площадь профильного поля ядра превышала значения цитоплазмы, поэтому с ЯЦК имел значения больше единицы, но у особей обеих групп значения ядерно-цитоплазматического коэффициента находятся примерно на одном уровне (таблица).
Сравнительные морфоцитохимические характеристики нейрон-глиальных популяций VI слоя крыши среднего мозга озерной лягушки (P. ridibundus), Мe [25; 75]
|
Показатель |
1 - группа P. ridibundus n = 10, m = 45,1 ± 5,0 г |
2 - группа P. ridibundus n = 7, m = 38,4 ± 4,2 г |
Значения р |
|
Параметры нейронов, мкм2 |
|||
|
Площадь профильного поля тела нейрона, Sт |
14,9 [12,9; 16,5] |
21,8 [18,8; 25,3] |
р < 0,001 |
|
Площадь профильного поля ядра, Sя |
9,3 [7,6; 10,6] |
13,0 [11,3; 15,2] |
р < 0,001 |
|
Площадь профильного поля цитоплазмы, Sц |
5,5 [5,0; 6,0] |
8,8 [7,8; 10,1] |
р < 0,001 |
|
Структурный ядерно-цитоплазматический коэффициент, сЯЦК |
1,6 [1,5; 1,8] |
1,5 [1,3; 1,7] |
р < 0,001 |
|
Показатели системы «нейрон-глия» (1 мм2), абс. число |
|||
|
Плотность нейронов |
6650,1 [5818,9; 8312,6] |
6580,8 [5911,2; 7758,5] |
р = 0,35 |
|
Плотность глиисвоб. |
3463,6 [2770,9; 4433,4] |
3048,0 [2632,3; 3602,1] |
р < 0,001 |
|
Плотность глиисат. |
2216,7 [1847,3; 2493,8] |
2147,4 [1773,4; 2493,8] |
р = 0,53 |
|
Глио-нейрональный индекс свободная глия, ГНИ своб. |
0,5 [0,4; 0,7] |
0,5 [0,4; 0,5] |
р < 0,001 |
|
Глио-нейрональный индекс сателлитнаяглия, ГНИ сат. |
0,3 [0,3; 0,4] |
0,3 [0,3; 0,4] |
р = 0,89 |
|
Оценка хроматофилии нейронов, перерасчет на 100 клеток, абс. число |
|||
|
Нормохромные нейроны |
65,0 [57,1; 72,2] |
70,6 [59,4; 76,2] |
р = 0,004 |
|
Гиперхромные нейроны |
23,3 [16,7; 30,0] |
17,0 [12,8; 22,2] |
р < 0,001 |
|
Гипохромные нейроны |
12,5 [8,7; 14,3] |
12,0 [10,3; 14,0] |
р = 0,39 |
|
Состояние фонда общих водонерастворимых белков, пкг/мкм2 |
|||
|
Концентрация белков в цитоплазме, Сц |
0,5 [0,5; 0,5] |
0,5 [0,4; 0,5] |
р < 0,001 |
|
Концентрация белков в ядре, Ся |
0,2 [0,2; 0,3] |
0,3 [0,2; 0,4] |
р < 0,001 |
|
Регуляторный ЯЦК (Ся/Сц), рЯЦК |
0,5 [0,3; 0,6] |
0,7 [0,5; 0,8] |
р < 0,001 |
|
Содержание белков в цитоплазме, Мц |
2,8 [2,3; 3,2] |
4,0 [3,5; 4,7] |
р < 0,001 |
|
Содержание белков в ядре, Мя |
2,0 [1,5; 2,5] |
3,9 [2,9; 5,0] |
р < 0,001 |
|
Функциональный ЯЦК (Мя/Мц), фЯЦК |
0,8 [0,6; 1,0] |
1,0 [0,7; 1,2] |
р < 0,001 |
Примечание: достоверность различий по сравнению с показателями особей пруда п. Шарыпово (критерий Манна – Уитни, р < 0,05).
Варьирование показателей профильного поля тел клеток, ядер и цитоплазмы нейронов VI слоя крыши среднего мозга у P. ridibundus, следует связать с активным распространением данного вида земноводных по территории южной части Красноярского края, и существованием в среде с комплексом разнообразных по действию экологических факторов, требующих для выживания переадаптации интегративной системы на всех уровнях организации, включая популяционно-клеточный. Выявленную вариабельность линейных параметров клеток можно рассматривать как вариант реактивности нейронов VI слоя крыши среднего мозга P. ridibundus, приспосабливающегося к водно-наземным условиям существования в биотопах с разным уровнем воздействия антропогенных факторов.
Плотность распределения нейронов в исследуемом слое у озерных лягушек различных биотопов имеет практически идентичные показатели. Значения плотности сателлитных глиоцитов VI слоя крыши среднего мозга у амфибий обеих групп также имеют близкие незначимые различия. Полученные данные плотности свободных глиоцитов выше плотности сателлитных глиоцитов у P. ridibundus обеих групп, что указывает на оптимизацию пролиферативных процессов и миграцию глиоцитов. Увеличение числа свободных глиоцитов у P. ridibundus группы 1 свидетельствует о включении компенсаторных механизмов, направленных на стабилизацию функционирования нейронов VI слоя крыши среднего мозга земноводных. Показатели ГНИ своб. преобладают над ГНИ сат. и значимо выше у лягушек 1 группы (таблица).
В популяциях нейронов VI слоя крыши среднего мозга по оценке хроматофильной субстанции у озерных лягушек 1 и 2 групп доминировал нормохромный тип клеток с небольшим содержанием гиперхромных нейроцитов и минимальным количеством гипохромных активно работающих клеток (таблица, рисунок).
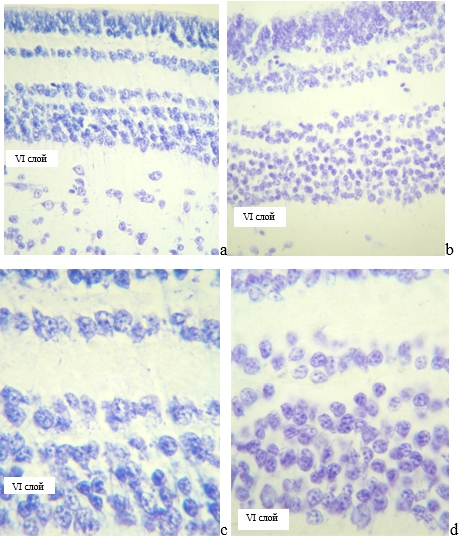
Крыша среднего мозга озерной лягушки, I – VI слои. Окраска по Нисслю
а, с – P. ridibundus пруд п. Шарыпово, b, d – P. ridibundus пойма р. Берешь Шарыповский район, ув. 10х40; с, d – нейроны VI слоя, ув. 10х100
У P. ridibundus группы 1 в нейрональной популяции VI слоя наблюдается сдвиг числа нейронов в сторону уменьшения нормохромных клеток и увеличения гиперхромных нейронов, накапливающих тигроид, что свидетельствует о подключении компенсаторных механизмов. Темные гиперхромные нейроны накапливают и не выводят за пределы клеток рибонуклеопротеидные комплексы, что указывает на интенсификацию адаптационных механизмов и создание запасающего фонда в клетках, как приспособление земноводных к обитанию в пруде п. Шарыпово.
В пределах популяций VI слоя крыши среднего мозга у амфибий изученного вида показатели белкового фонда (содержание и концентрация белков) в цитоплазме нейронов выше, чем в ядре. Данные внутриклеточные перестройки обеспечивают нормализацию метаболических процессов и адаптацию к воздействию факторов среды, и также рассматриваются как компенсаторные проявления на уровне клеточных популяций головного мозга.
Оценка состояния белкового фонда нейронов VI слоя крыши среднего мозга показала, что средние значения концентрации белков в цитоплазме клеток (Сц) в изученных биотопах имеют близкие значения показателей и превышают значения концентрации белков в ядре (Ся) в 2,5 и 1,7 раза у P. ridibundus 1 и 2 биотопов соответственно. Значения рЯЦК у особей обоих биотопов варьируют в пределах от 0,5 до 0,7, что обусловлено преобладанием концентрации плотных веществ в цитоплазме над таковыми в ядре, при более высоких линейных параметрах последних. При этом значения рЯЦК выше у особей группы 2 в 1,4 раза. Полученные данные также свидетельствуют о снижении интенсивности биосинтеза белка в ядрах клеток и пластического обмена в нейроцитах в целом.
Высокие показатели содержания белков в цитоплазме и ядрах клеток свидетельствуют о повышенной функциональной активности нейронов VI слоя у P. ridibundus обеих групп и преобладанием процессов биосинтеза белков над их использованием, что свидетельствует о компенсаторных процессах, направленных на восстановление запаса белков, необходимых для функционирования нейронов и передачи нервных импульсов. У P. ridibundus группы 1 содержание белков в цитоплазме нейронов (Мц) в 1,4 раза превышает их содержание в ядре (Мя). У лягушек группы 2 показатели Мц и Мя клеток имеют аналогично высокие и практически идентичные показатели, что свидетельствует о повышении пластического обмена и функциональной активности клеток изученного слоя. Высокие значения фЯЦК (0,8–1,0) у особей обоих биотопов также связаны с превышением содержания белков в цитоплазме нейронов по сравнению с ядром (таблица).
У амфибий группы 1 (с повышенным уровнем загрязнения) концентрация белков в цитоплазме высокая, в ядре низкая, при среднем содержании белков в цитоплазме и низком – в ядре. Данное сочетание связано с высокой функциональной нагрузкой нейронов у особей, испытывающих воздействие негативных факторов среды.
Заключение
Выполненный анализ морфоцитохимического состояния клеток VI слоя крыши среднего мозга P. ridibundus, обитающих на территории с повышенным уровнем загрязнения, выявил ряд однонаправленных компенсаторно-приспособительных изменений: наблюдается уменьшение линейных параметров нейронов, увеличение плотности распределения нервных клеток и глиоцитов, уменьшение числа нормохромных клеток и увеличение гиперхромных нейронов, снижение содержания в цитоплазме и ядрах нейронов общих водонерастворимых белков. Выявленные адаптивные перестройки нейрон-глиальных клеточных популяций более выражены у амфибий (группа 1) из популяции зоны малоэтажной застройки (п. Шарыпово), они направлены на поддержание гомеостаза в организме амфибий, в условиях, сочетающих урбанизацию и загрязнение.
Морфоцитохимические перестройки нейрон-глиальных популяций VI слоя крыши среднего мозга указывают на наличие в среде обитания P. ridibundus факторов, ведущих к включению компенсаторных механизмов, находящихся на границе нормы, и нарастанием в изученной клеточной популяции деструктивных процессов. Некоторая пластичность нейроморфологических характеристик нейронов головного мозга позволяет земноводным адаптироваться к антропогенным воздействиям среды, поддерживать жизнеспособность в трансформированных биотопах и распространяться на новые территории Красноярского края.