



Современный растительный покров степной зоны – поля различных агрофитоценозов, разделенные линиями искусственных лесополос. От нетронутой степи остались лишь небольшие участки. Чаще всего под степью сейчас понимают не целину, а залежь, т.е. участок земли, исключенный из хозяйственной деятельности человека. На таком поле сначала появляются только сорные растения, а затем с каждым годом в травостое все больше и больше начинают появляться растения, характерные для степи. Если участок будет предоставлен самому себе, постепенно он превратится в настоящую целину, практически неотличимую от девственной степи. Таким образом, протекает процесс естественного остепнения [2].
Результаты исследований ряда авторов показали, что на залежных участках в зависимости от времени остепнения угодий происходит смена растительных сообществ с увеличением видового разнообразия от сорных растений к типичным степным сообществам. Этот процесс влечет за собой увеличение численности всех исследуемых групп мелких членистоногих – почвенных клещей и ногохвосток [8,9].
По данным Н. В. Евсеевой [4], установлено изменение физических и химических свойств чернозема обыкновенного на разновозрастных залежах. Отмечено улучшение структурного состояния почв, повышение коэффициента структурности и водопрочности почвенных агрегатов (5-летняя – 3,04 и 0,92; 70-летняя – 3,46 и 1,44), изменяется качественный состав гумуса за счет увеличения содержания доли кислот и уменьшения доли негидролизуемого остатка. Содержание гумуса возрастает в ряду 5-; 15-; и 70-летних залежах (соответственно 2,68-2,74-3,40 %).
Это объясняется тем, что в деструкции органического вещества, скапливающегося в пахотном горизонте почвы, принимают участие различные группы мелких членистоногих, выполняющие роль регуляторов микробиологической активности, формирующие многозвеньевые сукцессионные ряды по мере минерализации субстрата и его дальнейшей гумификации.
В силу своих физических особенностей (высокий уровень смертности и быстрое наращивание численности) мелкие членистоногие наиболее чутко и быстро реагируют на изменение гидротермического и химического состава почв. Так, в летний период (июль) из-за высоких температур и низкой влажности почвы в пахотных горизонтах наблюдается резкое снижение численности микроартропод, а также видового состава и количества особей ногохвосток.
Таким образом, мелкие членистоногие (клещи, ногохвостки) вместе с микрофлорой ускоряют процесс минерализации органических остатков, что делает микроартропод важными почвообразователями [5].
Цель настоящих исследований – выявить закономерности сезонной динамики агрохимических показателей залежных участков каштановых почв юго-восточных районов Ростовской области и их влияние на почвенную биоту (клещей и ногохвосток).
Объекты и методы исследований
Район исследований относится к сухостепной зоне юга Европейской территории России и находится на равнинной территории западных склонов Южных Ергеней. Сухостепная зона только в Южном федеральном округе России занимает около 5575 тыс. га.
Каштановые почвы – основной зональный тип почв сухих степей – формируются под влиянием ксерофитной растительности, покров которой изреженный и низкорослый со степенью покрытия около 50–70 %. Для настоящих степей Приазовья запас фитомассы составляет 130–140 ц/га, в сухих степях – меньше 100 ц/га, а для сухих солонцеватых степей – 39-59 ц/га [8]. Ежегодный прирост также в 2,5–3 раза меньше, чем в настоящих степях. В структуре фитомассы преобладают подземные органы. Запасы мортмассы в степях близки к запасам фитомассы.
Формирование комплексности почвенного покрова – характерная черта почв сухих степей [2;3]. Основные причины комплексности: микрорельеф, различный характер увлажнения и солонцового режима, солонцеватость почв, вынос землероями на поверхность засоленного грунта и, как следствие, пятнистая неоднородность почв и растительности. Как правило, основные составляющие пятнистого комплекса следующие: каштановые обычные почвы, каштановые солонцы, лугово-каштановые почвы [2;3].
Каштановые почвы характеризуются следующими генетическими горизонтами:
А – гумусово-аккумулятивный горизонт каштанового цвета с сероватым или коричнево-сероватым оттенком. В естественном состоянии структура мелкозернисто-порошистая, часто с поверхности слоеватая. При распашке становится глыбистой и порошистой. Мощность горизонта 15–30 см.
АВ – гумусовый переходный горизонт, светлее предыдущего, серо-буроватый, каштановый, с признаками призмовидности. Обычно вскипает от НСl. Нижняя граница горизонта 45–60 см.
В – переходный горизонт, неоднородно-окрашенный с гумусированными пятнами и языками. Неоднородность окраски усиливается пятнами кротовин, гумусированными ходами червей и новообразованиями карбонатов. Мощность горизонта около 10 см.
Вca(Сca) – иллювиальный десуктивно-карбонатный горизонт, пропитан карбонатами кальция. Новообразования карбонатов выделяются в виде обильной белоглазки, прожилок или мучнистых скоплений. Нижняя граница горизонта прослеживается до глубины 100–150 см.
Bs(Cs) – иллювиальный горизонт скопления гипса и легкорастворимых солей. Выделения карбонатов редкие. Гипс в виде друз, гнезд, прожилок. В нижней части горизонта могут проявляться выделения легкорастворимых солей. Нижняя граница профиля расположена на глубине 180–250 см.
С – материнская порода различного генезиса.
Следовательно, тип каштановых почв определяют гумусовые горизонты А+АВ и солевые горизонты Вca + Bs. Мощность гумусового профиля всего около 50 см, профиль почвы в целом около 200 см.
Каштановые почвы в районе исследования относятся к восточно-европейской фации сухих степей, которые отличаются континентальным климатом с морозными зимами. Широко распространены солонцы и солонцеватые почвы с черно-полынной растительностью. Среди материнских пород часто встречаются засоленные глины разного генезиса. Профиль каштановых почв восточно-европейской фации находится в состоянии мерзлотного покоя в течение 2–3 месяцев [3]. В горизонте А (гумусово-аккумулятивный) каштановых почв содержание гумуса 2,7–2,8 %, общие запасы его по профилю 90–160 т/га. Значительная доля участия фульвокислот, если в верхней части гумусового горизонта гумус фульватно-гуматный, то в нижней – гуматно-фульватный [7].
По классификации 1977 года в типе каштановых почв выделяется 3 подтипа: светло-каштановые, каштановые и темно-каштановые почвы (табл. 1) [3].
Таблица 1
Подтиповые признаки каштановых почв
|
Подтип |
Глубина залегания новообразований, см |
Мощность А+В, см |
Содержание гумуса в гор. А, % |
|
|
CaCO3 |
CaSO4 |
|||
|
Темно-каштановые |
45–60 |
˃ 170 |
35–50 |
4–5 |
|
Каштановые |
40–45 |
150–170 |
30–40 |
3–4 |
|
Светло-каштановые |
35 |
˂ 150 |
25–35 |
2–3 |
В 2014 г. на территории 6 районов (Орловского, Зимовниковского, Мартыновского, Пролетарского, Дубовского, Заветинского) Юго-Востока Ростовской области для изучения агрохимических показателей были отобраны смешанные образцы почвы (0–20 см). Для изучения биоты металлической рамкой объемом 125 см3 в 15-кратной повторности были отобраны почвенные образцы на глубину 0–20 см. Экстракция микроартропод проводилась по методике Балога (1958) [10] без электрического обогрева в течение 7 дней. Разбивка на группы и подсчет проводились под бинокуляром МБС-1. Гумус определяли по методу Тюрина (в модификации Симакова) нитратный азот – ионометрическим методом, аммиачный азот – фотоколориметрически с реактивом Несслера, рН – потенциометрическим методом, подвижный фосфор и калий – по методу Мачигина [1, 6].
Результаты и их обсуждение. Почвы изучаемых районов представлены тремя подтипами каштановых почв: темно-каштановые – Мартыновский и Пролетарский районы, каштановые – Орловский район, светло-каштановые почвы – Зимовниковский, Заветинский и Дубовский районы. Содержание гумуса в почвенных образцах соответствует подтиповым признакам этих почв (табл. 2). В мае максимальное содержание гумуса 5,42 и 3,60 % отмечено в темно-каштановых почвах Мартыновского и Пролетарского районов, минимальное в светло-каштановых почвах Заветинского и Зимовниковского районов – 2,50 и 2,15 % соответственно.
Таблица 2
Агрохимические показатели залежных участков каштановых почв (0–20 см), май, 2014 г.
|
№ обр |
Район исследования, почва |
рН |
Гумус, % |
NH4+, мг/100г |
NO3 -, мг/100г |
Р2О5, мг/100г |
К2О, мг/100г |
|
1 |
Мартыновский р-н, темно-кашт |
8,12 |
5,42 |
9,8 |
4,70 |
8,20 |
21,01 |
|
2 |
Пролетарский р-н, темно-кашт. |
7,93 |
3,60 |
5,50 |
3,90 |
6,00 |
17,11 |
|
3 |
Орловский р-н, каштановая |
8,19 |
3,11 |
4,20 |
3,40 |
4,60 |
14,39 |
|
4 |
Зимовниковский р-н св.-каштан. |
8,35 |
2,15 |
3,50 |
2,90 |
3,40 |
17,98 |
|
5 |
Дубовский р-н, св.-каштановая |
8,33 |
3,00 |
2,62 |
3,00 |
3,80 |
15,20 |
|
6 |
Заветинский р-н, св-каштановая |
8,38 |
2,50 |
1,78 |
3,15 |
3,90 |
16,10 |
Таблица 3
Агрохимические показатели залежных участков каштановых почв (0–20 см), сентябрь, 2014 г.
|
№ обр |
Район исследования, |
рН |
Гумус, % |
NH4+, мг/100г |
NO3 -, мг/100г |
Р2О5, мг/100г |
К2О, мг/100г |
|
1 |
Мартыновский р-н, темно-кашт |
8,00 |
5,60 |
2,90 |
2,10 |
6,80 |
20,00 |
|
2 |
Пролетарский р-н, темно-кашт. |
7,74 |
3,70 |
2,40 |
0,15 |
4,50 |
15,32 |
|
3 |
Орловский р-н, каштановая |
8,05 |
3,30 |
2,20 |
0,12 |
3,20 |
12,56 |
|
4 |
Зимовниковский р-н св.-каштан. |
8,20 |
2,32 |
1,90 |
0,25 |
2,40 |
13,80 |
|
5 |
Дубовский р-н, св.-каштановая (пастбище, целина) |
8,22 |
3,15 |
1,20 |
0,22 |
2,60 |
12,60 |
|
6 |
Заветинский р-н, св-каштановая (пастбище, целина) |
8,25 |
2,80 |
0,90 |
0,25 |
2,50 |
13,00 |
К началу осени содержание гумуса во всех почвах увеличивается в среднем на 4 % (2–6 %), что связано с поступлением и накоплением органического вещества.
Количество подвижных форм NPK в темно-каштановых почвах выше, чем в каштановых и светло-каштановых почвах, что обусловлено большим содержанием гумуса.
Содержание обменного калия в изучаемых каштановых почвах характеризуется как повышенное и остается стабильным и осенью.
Высокой обеспеченностью Р2О5 характеризуются все исследуемые каштановые почвы весной. К осени – в каштановой и светло-каштановой почве степень обеспеченности подвижным фосфором снижается до средней.
По содержанию NH4+ и NO3 также прослеживается снижение от высокой обеспеченности до низкой, что обусловлено снижением биологической активности.
Таким образом, содержание NH4+ , NO3-, Р2О5 и К2О в каштановых почвах к осени уменьшается, что объясняется снижением биологической активности (табл. 2,3) в связи с высокими температурами воздуха и небольшим количеством осадков (рис. 1, 2). На рис. 1 и 2 приведены данные по температуре воздуха и количеству осадков на примере пос. Заветное Ростовской области.
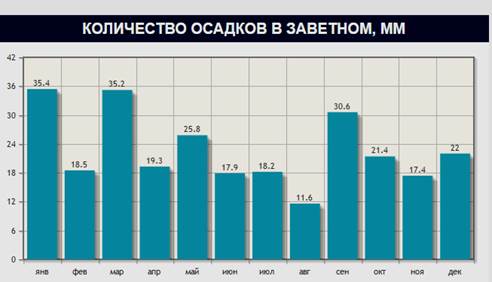
Рис.1. Количество осадков в пос. Заветное Ростовской области, мм, 2014 г.
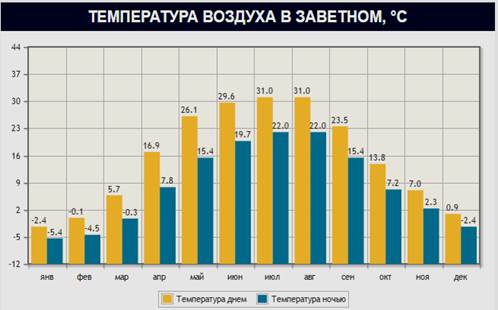
Рис. 2. Температура воздуха в пос. Заветное Ростовской области, ˚С, 2014 г.
Также установлено, что к осени происходит снижение pH почв (табл. 3) в среднем на 0,15 единиц рН (0,11–0,19), что составляет в среднем около 2 %. Уменьшение рН вероятно связано с поступлением органического вещества, в состав которого входят органические кислоты. Также в почвенный раствор продолжает поступать СО2 – продукт метаболизма почвенной фауны, микробозеноза и корневой системы растений. В результате проведенных исследований было выявлено, что наибольшая численность микроартропод (тыс. экз./м2) наблюдалась в мае – в залежах на территории Пролетарского и Мартыновского районов – 68,5 и 60,2, в том числе панцирных клещей – 22,3 и 9,7, гамазовых – 28,0 и 30,2, клещей акароидно-тромбидиформного комплекса – 1,5 и 0,7, ногохвосток – 15,2 и 17,1, прочих беспозвоночных – 1,5 и 2,5 (табл. 4).
Таблица 4
Численность микроартропод (тыс. экз./м2) каштановых залежных почв в юго-восточных районах Ростовской области (май 2014 г.)
|
Группы микроартропод |
Марты-новский р-н |
Пролетар-ский р-н |
Орловский р-н |
Зимовни-ковский р-н |
Дубовский р-н |
Заветин-ский р-н |
|
Панцирные клещи |
9,7±0,2 |
22,3±0,1 |
12,5± 0,6 |
20,5±0,82 |
8,2±0,5 |
7,5±0,3 |
|
Гамазовые клещи |
30,2±0,5 |
28,0±0,6 |
15,4±0,9 |
10,1±0,3 |
10,1±0,4 |
11,3±0,6 |
|
Акароидно-тромбидиформный комплекс клещей |
0,7±0,7 |
1,5±0,4 |
1,3±0,4 |
1,6±0,3 |
0,5±0,3 |
0,3±0,2 |
|
Ногохвостки |
17,1±0,2 |
15,2±0,1 |
7,3±0,2 |
3,5±0,5 |
3,4±0,6 |
4,5±0,5 |
|
Прочие беспозвоночные |
2,5±0,7 |
1,5±0,6 |
1,0 ±0,5 |
0,4±0,8 |
0,5±0,9 |
0,7±0,6 |
|
Всего микроартропод |
60,2±1,3 |
68,5±1,7 |
37,5±1,5 |
36,1±1,2 |
22,7±0,9 |
24,3±1,0 |
Исследование вертикального распределения микроартропод по почвенному профилю на глубину 0–20 см показало, что основная масса мелких членистоногих (> 60 %) была сосредоточена в горизонтах 0–10 см в течение вегетационного периода. Это объясняется тем, что в этом почвенном горизонте сосредоточена основная масса корневых систем многолетних трав.
В осенний период отмечено постепенное снижение численности панцирных клещей и клещей акароидно-тромбидиформного комплекса с увеличением глубины. Максимальная численность у гамазовых клещей наблюдалась в слое 10–15 см. Для ногохвосток и прочих беспозвоночных отмечен всплеск численности в слое 0–5 см.
К осени (сентябрь) численность микроартропод снижалась. Наибольшая численность микроартропод (тыс. экз./м2) наблюдалась так же, как и весной, в залежах на территории Пролетарского и Мартыновского районов – 50,0 и 49,9, в том числе панцирных клещей – 11,3 и 7,3, гамазовых – 26,1 и 25,1, клещей акароидно-тромбидиформного комплекса – 1,5 и 0,8, ногохвосток – 10,1 и 14,6, прочих беспозвоночных – 1,0 и 2,1 (табл. 5).
Таблица 5
Численность микроартропод (тыс. экз./м2) каштановых залежных почв в юго-восточных районах Ростовской области (сентябрь 2014 г.)
|
Группы микроартропод |
Марты-новский р-н |
Пролетар-ский р-н |
Орловский р-н |
Зимовни-ковский р-н |
Дубов- ский р-н |
Заветинский р-н |
|
Панцирные клещи |
7,3±0,5 |
11,3±0,3 |
9,2± 0,8 |
10,1±0,2 |
6,2±0,6 |
6,0±0,2 |
|
Гамазовые клещи |
25,1±0,4 |
26,1±0,7 |
12,7±0,5 |
9,3±0,2 |
9,1±0,8 |
8,9±0,2 |
|
Акароидно-тромбидиформный комплекс клещей |
0,8±0,9 |
1,5±0,7 |
1,2±0,7 |
1,5±0,7 |
0,5±0,4 |
0,5±0,3 |
|
Ногохвостки |
14,6±0,4 |
10,1±0,5 |
4,2±0,5 |
1,3±0,1 |
2,2±0,3 |
1,9±0,6 |
|
Прочие беспозвоночные |
2,1±0,3 |
1,0±0,5 |
1,0 ±0,5 |
0,3±0,6 |
0,5±0,2 |
0,6±0,2 |
|
Всего микроартропод: |
49,9±1,2 |
50,0±1,5 |
28,3±2,6 |
22,5±1,5 |
18,5±2,2 |
17,9±1,7 |
В силу своих физических особенностей (высокий уровень смертности и быстрое наращивание численности) мелкие членистоногие наиболее чутко и быстро реагируют на изменение гидротермического и химического состава почв [5]. Так, в осенний период (сентябрь) наблюдается резкое снижение численности микроартропод, а также количества особей ногохвосток, из-за высоких температур и низкой влажности почвы в пахотных горизонтах [8,9].
Выводы
К осени снижается рН, а содержание гумуса несколько увеличивается, что связано с поступлением органики. Численность биоты снижается осенью в почве исследуемых районов. Следовательно, снижение биологической активности, скорее всего, связано с климатическими показателями – температурой и осадками. Количество микроартропод коррелирует прямо пропорционально с содержанием гумуса и обратно пропорционально уровню рН. С увеличением щелочности почв (рН ˃8,20) их численность достоверно снижается. Содержание минерального азота (NH4+ и NO3–) к осени уменьшается, что тоже объясняется снижением биологической активности и климатическими показателями.
Исследования выполнены в рамках базовой части внутреннего гранта ЮФУ по проекту 213.01-2015/003ВГ.