



Цель исследования
Изучить морфологические и ультраструктурные изменения нейронов внутреннего ядерного слоя сетчатки крыс при воздействии света высокой интенсивности в динамике.
Материал и методы исследования
Изучены сетчатки 40 беспородных половозрелых белых крыс-самцов. На крыс 1-й группы (n=20) воздействовали в течение 6 часов люминесцентными лампами ЛБ-40, имеющими максимум излучения в желто-зеленой области спектра. Для освещения использовалась специальная установка из прямоугольных рефлекторов с вмонтированными в них лампами, освещающая клетку с 5 сторон. Освещенность животных составляла 6000 лк. В качестве контрольной группы использовали интактных крыс (n=20). Все животные содержались в условиях искусственного светового режима: 12 ч день, 12 ч ночь, с интенсивностью дневного освещения 25 лк. Декапитацию производили под эфирным наркозом через 1, 7, 14, 30 сут после светового воздействия, в аналогичные сроки производили взятие контрольного материала.
Для электронной микроскопии центральные участки задней стенки глаза фиксировали в растворе, содержащем смесь 4%-ного параформальдегида и 0,5%-ного глютаральдегида на 0,2 М какодилатном буфере (рН 7,4). Материал постфиксировали в 2%-ном растворе четырехокиси осмия и заливали в эпон. Полутонкие срезы окрашивали толуидиновым синим, ультратонкие - контрастировали уранилацетатом и цитратом свинца, просматривали и фотографировали в электронном микроскопе JEM–7А. На полутонких срезах сетчатки производили подсчет пикноморфных биполярных, амакринных и горизонтальных ассоциативных нейронов на 200 соответствующих клеток. На электронных микрофотографиях внутреннего ядерного слоя определяли удельную площадь органелл ассоциативных нейронов. Измерения проводили с помощью открытой квадратной тестовой решетки с шагом 5 мм при увеличении 10000.
Статистическую обработку результатов проводили методами вариационной статистики. Для оценки достоверности различий при сравнении средних величин использовали непараметрический критерий Манна-Уитни.
Результаты исследования и их обсуждение
На 1-е сутки после светового воздействия наблюдается достоверный рост процентного содержания пикноморфных ассоциативных нейронов (рис. 1), а также клеток со светлым типом деструкции, на ультраструктурном уровне характеризующийся снижением содержания органелл. Так, удельная площадь митохондрий в таких клетках снижалась в 1,5 раза, гранулярной эндоплазматической сети в 2,3 раза, комплекса Гольджи в 1,4 раза (контроль 15,58±2,08%; 26,79±2,41%; 2,71±0,13% соответственно; р<0,05). Максимум деструктивных изменений в нейронах внутреннего ядерного слоя зарегистрирован к 7-м суткам после освещения, когда процентное содержание пикноморфных нейронов возрастает относительно контрольного значения до 21,00±1,29% (контроль 1,85±0,13%; p<0.05; рис. 1). Пик деструкции ассоциативных нейронов совпадает с очаговым выпадением наружных слоев сетчатки вследствие массовой гибели нейросенсорных клеток и пигментоцитов [2; 5].
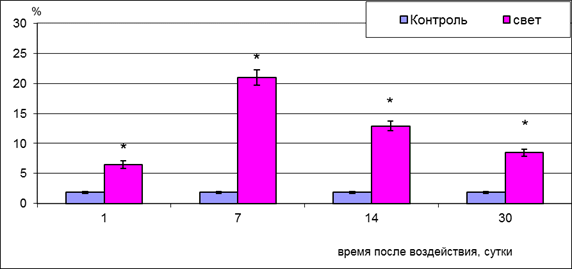
Рис. 1. Процентное содержание пикноморфных нейронов внутреннего ядерного слоя сетчатки крыс при световом воздействии
Примечание: статистически достоверные отличия (p<0.05) отмечены * - при сравнении с контролем.
Экспериментальное воздействие высокоинтенсивного света на сетчатку приводит к глубокому обесцвечиванию ретиналя и развитию процессов окисления в наружных сегментах и мембранах органелл внутренних сегментов фоторецепторов. При этом пигментоциты, находясь в тесном контакте с наружными сегментами нейросенсорных клеток, подвергаются мощной атаке свободными радикалами и при повреждении сами становятся индукторами активных форм кислорода [4]. Вероятно, срыв механизмов антиоксидантной защиты приводит к распространению свободных радикалов и продуктов пероксидации во внутренние слои сетчатки, что вызывает повреждение мембранных органелл, восприимчивых к окислению.
Активизация свободнорадикального окисления и перекисного окисления липидов вызывает также деструкцию высокомембранных структур синаптических контактов наружного сетчатого слоя. В отдельных участках сетчатки он полностью отсутствует, что прерывает трехнейронную цепь и опосредованно вызывает деструкцию нейронов внутреннего ядерного слоя. Деструкция нейронов II порядка во многом зависит от степени поражения нейросенсорных клеток и объясняется потерей синаптического входа от них и невозможностью выхода трофических факторов и медиаторов [7; 10]. На 14-30-е сутки количество пикноморфных нейронов снижается до 12,9±0,79% и 8,45±0,56% соответственно (контроль 1,85±0,13%; p<0.05). Данная динамика, возможно, связана с фагоцитозом деструктивных нейронов радиальной глией, о чем косвенно можно судить по наличию в цитоплазме радиальных глиоцитов остаточных телец (рис. 2). В эти сроки внутренний ядерный слой представлен клетками со светлым типом деструкции, а также гипертрофированными отростками радиальной глии (рис. 3), вероятно, осуществляющими защиту сохранившихся нейронов.
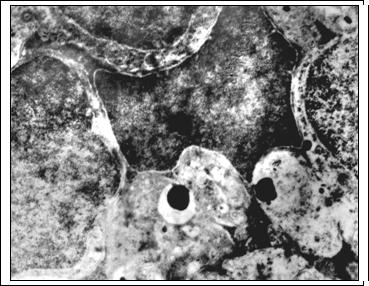
Рис. 2. Остаточные тельца в цитоплазме радиального глиоцита через 14 суток после светового воздействия. Я - ядро радиального глиоцита; О - остаточные тельца. Ув. 6500
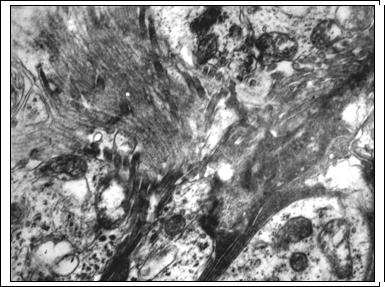
Рис. 3. Глиальная пролиферация во внутреннем ядерном слое через 14 суток после освещения. ОРГ - отростки радиальной глии; А - ассоциативные нейроны. Ув. 8000
Реакция ассоциативных нейронов и радиальных глиоцитов в ответ на световое воздействие в целом схожа с процессами, происходящими в сетчатке при различных нейродегенеративных состояниях. Процесс ремоделирования сетчатки напоминает ответ центральной нервной системы на деафферентацию или травму и включает, главным образом, гибель нейронов, миграцию глиальных популяций и неосинаптогенез [7]. Обращает на себя внимание тот факт, что распределение радиальных глиоцитов во внутреннем ядерном слое происходит неравномерно, основная часть их ядер сконцентрирована у витреального края этого слоя и находится в непосредственном контакте с телами амакринных нейронов. Отмечено, что большая часть этих глиоцитов при световом воздействии подвержена деструкции. Как известно, глия обеспечивает нормальное функционирование нейронов, а ее гибель, косвенно, инициирует деструкцию нейронов. Так, например, радиальные глиоциты участвуют в метаболизме нейротрансмиттера глутамата, который в больших концентрациях обладает нейротоксичностью и индуцирует апоптотическую гибель нейронов [3; 9].
Среди ассоциативных нейронов наиболее подвержены деструкции амакринные нейроны и наименее - горизонтальные (рис. 4). Так, количество пикноморфных амакринных нейронов на 7-е сут после светового воздействия достигает 26,6±1,26%, биполярных 17,4±0,69% и горизонтальных 13,6±1,24%, что значимо отличается от таковых в контроле (1,05±0,12%, 0,8±0,08%, 0% соответственно p<0.05).
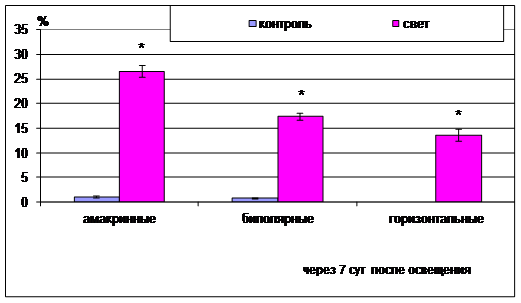
Рис. 4. Процентное содержание пикноморфных ассоциативных нейронов (амакринных, биполярных и горизонтальных) при высокоинтенсивном световом воздействии
Выраженные морфологические и функциональные изменения амакринных и биполярных нейронов с сокращением числа дендритов, прорастанием аксонов, потерей нейронами глутаматных рецепторов развиваются также и при ряде других нейродегенеративных заболеваний сетчатки. Деструкция нейросенсорных клеток на начальном этапе развития заболеваний приводит к увеличению синаптической активности в биполярных и амакринных нейронах, следствием чего является истощение и перераспределение пула нейромедиаторов [6]. Вероятно, именно увеличение функциональной активности амакринных и биполярных нейронов лежит в основе большего их повреждения при воздействии света высокой интенсивности.
Выводы
Таким образом, при световом воздействии наиболее подвержены деструктивным изменениям амакринные и наименее – горизонтальные нейроны внутреннего ядерного слоя. Максимальное содержание пикноморфных нейронов наблюдается на 7-е сутки после воздействия, одновременно с очаговым выпадением наружных слоев сетчатки. В поздние сроки содержание деструктивно измененных нейронов достоверно снижается вследствие их фагоцитоза радиальными глиоцитами.
Рецензенты:
Солонский А.В., д.м.н., руководитель лаборатории нейробиологии ФГБУ «Научно-исследовательский институт психического здоровья» СО РАМН, г. Томск.
Мустафина Л.Р., д.м.н., профессор кафедры гистологии, эмбриологии и цитологии ГОУ ВПО «Сибирский государственный медицинский университет» Минздрава России, г. Томск.