



Поджелудочная железа, благодаря гормональной функции, принимает участие в формировании адаптивных реакций не только к изменяющимся параметрам внешней среды, но и является звеном общего адаптивного комплекса организма, который реагирует на изменение гомеостаза. Локальные изменения уровня инсулярных гормонов могут быть морфологической основой изменений экзокринной части поджелудочной железы [2, 7].
Для млекопитающих животных характерно наличие четырех типов эндокриноцитов, присутствующих в панкреатических островках поджелудочной железы и синтезирующих гормоны. Самые многочисленные клетки – это β-клетки или инсулиноциты, синтезирующие инсулин, значительно меньшее количество занимают α-клетки или глюкагоноциты, синтезирующие глюкагон, в единичном количестве присутствуют δ – клетки или соматостатиноциты синтезирующие соматостатин и РР-клетки синтезирующие панкреатический полипептид. Имеются также единичные сведения о популяциях гормонпродуцирующих клеток в экзокринной части поджелудочной железы [1, 5].
Использование иммуногистохимических методов выявления цитоархитектуры и видового состава эндокриноцитов, их соотношения в экзокринной и эндокринной частях поджелудочной железы таких млекопитающих как человек, приматы, мыши, свиньи, нутрии позволило выявить не только межвидовые, но и возрастные их различия у особей [4, 10, 11]. Сведения структурного строения, клеточного состава эндокриноцитов поджелудочной железы крупного рогатого скота в настоящее время отсутствуют.
Цель настоящей работы – иммуногистохимическое исследование структурного строения, клеточного состава и количественных показателей эндокриноцитов панкреатических островков, а также выявление и изучение эндокриноцитов и их скоплений, диффузно распределенных в экзокринной части поджелудочной железы крупного рогатого скота в постнатальном онтогенезе.
Материал и методы. В работе исследована поджелудочная железа крупного рогатого скота айширской породы. Для исследований было отобрано 15 голов клинически здоровых животных в разные периоды постнатального онтогенеза: 1-суточные, 1-, 3-, 6-месячные и в возрасте 1 и 3 года (по 3 животных в каждой возрастной группе). Убой животных проводился в соответствии с Директивой 2010/63/EU ЕВРОПЕЙСКОГО ПАРЛАМЕНТА И СОВЕТА ЕВРОПЕЙСКОГО СОЮЗА по охране животных, используемых в научных целях.
У каждого животного проводили отбор целой поджелудочной железы, которую фиксировали в 10% нейтральном забуференном формалине. В области головки железы вырезали кусочки размером 1см3. Материал проводили с использованием гистологического процессора замкнутого типа Tissue-Tek VIP™ 5 Jr (Sakura (Япония)), а затем заливали в гистологическую среду «Гистомикс» (БиоВитрум (Россия)) на станции парафиновой заливки Tissue-Tek® TEC™ 5 (Sakura (Япония)). Из полученных блоков делали гистологические срезы толщиной 5 - 7 мкм.
Для иммуногистохимических реакций использовали антитела к гормонам поджелудочной железы: моноклональные мышиные антитела к инсулину клон К36аС10 (Diagnostic BioSystems, Нидерланды 1:25 – 1:50), поликлональные кроличьи антитела к глюкагону (SpringBioScience, США), поликлональные кроличьи антитела к соматостатину (MONOSAN, Нидерланды) и кроличьи поликлональные антитела к панкреатическому полипептиду (DCS, Германия 1:25 – 1:50). Негативным контролем служили реакции с заменой первых антител раствором для разведения (SpringBioScience, США).
При проведении иммуногистохимических реакций депарафинированные и гидратированные срезы обрабатывали 3% раствором Н2О2 в течение 10 мин. для блокирования эндогенной пероксидазы. Затем срезы подвергали высокотемпературной обработке в 0,01М цитратном буфере (рН 6,0) в течение 40 мин. Инкубация срезов с первичными антителами на инсулин, глюкагон, соматостатин и панкреатический полипептид проводили во влажной камере при температуре 27ºС в течение 24 часов. Со вторыми козьими антителами инкубацию проводили в течени 60 минут во влажной камере при температуре 27ºС. После инкубации применяли высокочувствительную систему визуализации Reveal biotin-free polyvalent DAB (SpringBioScience, США). Микроскопию срезов проводили на световом микроскопе Olympus BX45 со встроенным фотоаппаратом.
С каждого препарата поджелудочной железы окрашенного позитивно на гормоны поджелудочной железы, выполняли по 10 цифровых снимков (в формате jpg, размером 3136×2352 пикселей в палитре 24 бит) случайно выбранных полей зрения при увеличении х100, х200, х400 и х1000. Визуально при увеличении объектива х200 проводили подсчет позитивно окрашенных островков, и их клеток – β-, α-, δ-, и РР- при увеличении х1000.
Результаты исследований. У 1-суточных телят при иммуногистохимических исследованиях при увеличении х200 в дольке железы насчитывается от 5 до 13 панкреатических островков различных размеров, в которых клетки имеют позитивную реакцию на инсулин, глюкагон, соматостатин и панкреатический полипептид.
β-эндокриноциты в островке образуют клеточные скопления и расположены в основном по периферии в виде незамкнутых колец. Количество их в островке составляет от 4 до 22. Одновременно визуализируется значительное количество островков состоящих полностью их β-эндокриноцитов, позитивная реакция на глюкагон, соматостатин и панкреатический полипептид в них отсутствовала. Количество α-эндокриноцитов в островке составляет от 1 до 19 клеток. Они расположены диффузно между β-эндокриноцитами по периферии островка, но сосредоточены только на одном из его полюсов. δ-эндокриноциты расположены на границе с кровеносными капиллярами, оплетающими островок, в виде замкнутой цепочки клеток, лежащих в один ряд. Количество клеток варьирует от 2 до 19. PP-эндокриноциты расположены в островке диффузно в центральной его части. Их количество немногочисленное и составляет от 1 до 3 клеток.
Кроме островков β-, δ-, и РР-эндокриноциты были отмечены и в экзокринной части железы. β-, и РР-эндокриноциты встречаются как одиночные, так и в количестве от 2 до 4. Визуализируются чаще всего они в междольковых выводных протоках между эпителиоцитами и среди панкреатоцитов ацинусов экзокринной части железы. δ-клетки чаще одиночные и наблюдаются между панкреатоцитами ацинусов.
В 1-месячном возрасте у телят количество эндокринных островков остается на прежнем уровне и составляет от 5 до 13. Отмечается появление островков в которых регистрируется перемещение скоплений β-эндокриноцитов от периферии в центральную часть. Имеются единичные островки уже с центральным расположением инсулин позитивных клеток. Количество β-эндокриноцитов в этом возрасте увеличивается и составляет от 10 до 26 клеток. α-эндокриноциты не претерпевают структурных перемещений по отношению к островку, количественный их состав остается на прежнем уровне. δ-эндокриноциты в количественном составе снижаются и составляют от 5 до 10 клеток. Располагаются они преимущественно в центре островка в виде скоплений. Количество РР-эндокриноцитов в этом возрасте увеличивается и равно от 3 до 7, расположение клеток остается на прежнем уровне в центре островка.
Наряду с вышеописанным продолжают обнаруживаться единичные небольшие кластеры и одиночные клетки, лежащие в экзокринной части железы и состоящие из β-, и РР-эндокриноцитов. δ-эндокриноциты в 1-месячном возрасте телят вне островков выявлены не были. Отмечается появление одиночных и редких α-эндокриноцитов между ацинусами панкреатоцитов.
В 3-месячном и 6-месячном возрастах количество эндокринных островков в дольках резко снижается и составляет от 2 до 6. Визуализируются островки, в которых β-эндокриноциты расположены в основном в центральной части или смещены на один полюс островка. В 3-месячном возрасте количество инсулин позитивных клеток составляет от 3 до 41, а в 6-месячном – от 5 до 34. α-эндокриноциты продолжают располагаться диффузно на периферии островка, занимая один из его полюсов. Количество их в 3-месячном возрасте равно от 4 до 20, в 6-месячном от 4 до 22. δ-эндокриноциты в обеих возрастных группах составляют от 2 до 15 клеток в 3-месячном возрасте и от 2 до 13 в 6-месячном. Расположены они в основном диффузно по всему периметру островка, однако встречаются единичные островки, в которых клетки лежат поодиночке, повторяя рисунок расположения у 1-суточных телят. РР-эндокриноциты в 3-месячном возрасте составляют от 3 до 10 клеток, в 6-месячном – от 4 до 8. Отмечается, что единичные клетки в 6-месячном возрасте из центра переходят на периферию островка и находятся на границе с экзокринной частью и кровеносными капиллярами, оплетающими эндокринный островок.
В экзокринной части железы у телят в 3-месячном и 6-месячном возрасте наблюдаются единичные β-, α-, δ- и РР-эндокриноциты, лежащие между панкреатическими ацинусами. Единичные β- и РР-эндокриноциты иногда просматриваются вокруг междольковых выводных протоков.
В возрасте 1 года у крупного рогатого скота количество островков уменьшается и составляет в дольке железы от 1 до 2, к 3-летнему возрасту количество островков увеличивается и равно от 1 до 7. β-эндокриноциты в островке расположены диффузно и занимают периферическое положение. Количество инсулин позитивных клеток в годовалом возрасте составляет от 4 до 24 клеток, а к 3-летнему возрасту увеличивается от 7 до 39. Количественный состав α-, и δ-эндокриноцитов по сравнению с вышеописанными возрастами существенно снижается, но остается на одном уровне от 1-года до 3-лет. Так, количество α-эндокриноцитов в годовалом возрасте составляет от 4 до 10 клеток, δ-эндокриноцитов – от 1 до 5, в 3-летнем возрасте – от 4 до 7 и от 1 до 3 соответственно. α-эндокриноциты лежат у одного из полюсов островка. δ-эндокриноциты расположены поодиночке, занимают центральное положение в островке. Просматриваются островки с одиночными соматостатин позитивными клетками, которые расположены на периферии островка. Количество РР-эндокриноцитов от годовалого возраста до 3-лет существенно уменьшается и составляет от 1 до 3. Расположены клетки диффузно. Встречаются островки, в которых клетки лежат в зоне перехода эндокринной части в экзокринную (Рис. 1, 2).
В экзокринной части железы в годовалом и 3-летнем возрасте встречаются только β-, δ- и РР-эндокриноциты, которые лежат поодиночке между панкреатическими ацинусами.
Обсуждение полученных данных. При иммуногистохимическом исследовании поджелудочной железы крупного рогатого скота от 1-суточного до 3-летнего возраста нами были выявлены некоторые особенности цитоархитектоники эндокриноцитов железы.
По данным Весниной И. А. (2001), Пузырева А. А. (2003) в поджелудочной железе у различных представителей позвоночных эндокринные островки состоят из 4 типов эндокриноцитов – β-, α-, δ- и РР-. Основным цитотипом являются инсулинобразующие клетки. Результаты наших исследований наглядно демонстрируют наличие вышеописанных клеток у крупного рогатого скота в составе эндокринных островков поджелудочной железы. Наибольшее количество в островке составляют β-эндокриноциты, содержание которых в 1-суточном возрасте составляет 35%, а к 3 годам увеличивается до 75%. Вторыми по количественному составу являются α- и δ-эндокриноциты, которые в 1-суточном возрасте составляют по 30%. Однако к 3-летнему возрасту количество их уменьшается. Так, содержание α-эндокриноцитов уменьшается до 13,4%, а δ-эндокриноцитов – до 5,8% от общего содержания клеток в островке. Самыми малочисленными клетками являются РР-эндокриноциты, количество которых в 1-суточном возрасте составляет 4,6% и 5,8% в 3-летнем возрасте крупного рогатого скота. Увеличение количества β-эндокриноцитов в постнатальном онтогенезе наблюдала Можейко Л. А. (2004) у кроликов. По ее данным, клеточный состав островков претерпевает наиболее сложную трансформацию, особенно увеличивается содержание β-клеток, что связано с меняющимися адаптивными условиями существования и питания, когда происходит совершенствование существующих структур для реализации возможности полноценного функционирования органа в качественно новых условиях. Относительно сведений о количественных изменениях α-, δ- и ΡΡ-эндокриноцитов в постнатальном онтогенезе у животных и человека мы в доступной литературе не нашли. По нашему мнению, снижение α- и δ-эндокриноцитов у крупного рогатого скота к 3-летнему возрасту связано с их угнетающими эффектами на экзокринную часть железы, что приводит к уменьшению выработки ферментов, снижению расщепления белков и всасыванию аминокислот в кишечнике. Увеличение количества PP-эндокриноцитов связано с их функцией – регуляцией выработки панкреатического сока, а также фермента амилазы.
По цитоархитектонике, основным типом островков у крупного рогатого скота до 1 года постнатального онтогенеза является плащевой. В них по расположению клеток выделяются зоны: гемоцеллюлярная (центральная) в которой лежат β- и ΡΡ-эндокриноциты и гетероцеллюлярная (периферическая) содержащая α- и δ-эндокриноциты. Однако в 3 года отмечается структурная перепланировка клеток. Из гемоцеллюлярной зоны β-эндокриноциты перемещаются в гетероцеллюлярную, δ- и ΡΡ-эндокриноциты просматриваются как в центральной, так и в периферической части островков, α-эндокриноциты не меняют своего местоположения. Вышеописанные изменения полюсного расположения β- и α-эндокриноцитов в островке характеризуют их как плащевых биполярных. Из литературных источников известно, что плащевой тип эндокринных островков встречается у грызунов и кроликов Можейко Л. А. (2004, 2011), причем расположение β-эндокриноцитов центральное, сгруппированы они в инсулиновое ядро. У человека и приматов расположение эндокриноцитов мозаичное, однако Ульяновская С. А (2013), Bocian-Sobkowska J. (1999), Кривова Ю. С. (2009), изучая развитие и цитодифференцировку островков Лангерханса у плодов человека, выявили, что островки с 20 недели развития плода относятся к биполярному типу. Начиная с 34-недели развития α- и β-клетки меняют свою структурную организацию. Происходит их перераспределение и формирование островков мозаичного типа.
Таким образом, изменения организации эндокринных островков на различных этапах постнатального онтогенеза могут рассматриваться как адаптивные в связи с совершенствованием существующих структур для реализации возможности полноценного функционирования органа. Изучение механизмов становления в постнатальном онтогенезе эндокринного аппарата поджелудочной железы – одного из звеньев эндокринной системы – имеет общебиологическое значение, так как дополняет имеющиеся сведения об ее уникальных свойствах.
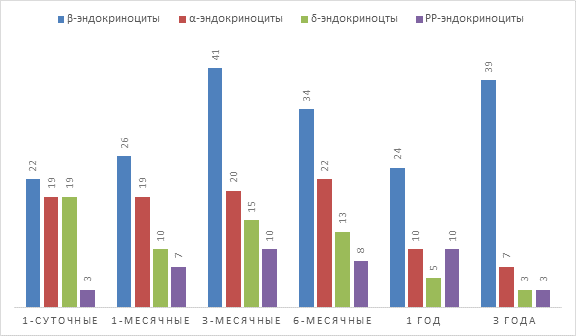
Рис. 1. Количественный состав эндокриноцитов в эндокринных островках крупного рогатого скота в постнатальном онтогенезе
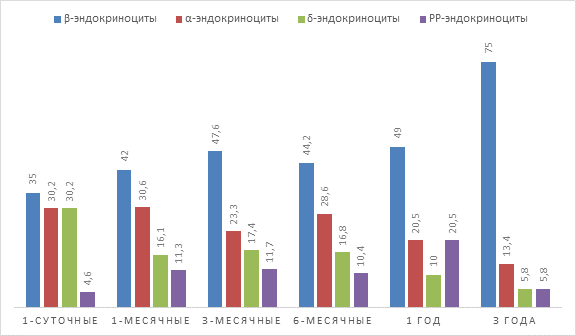
Рис. 2. Процентное соотношение эндокриноцитов в эндокринных островках крупного рогатого скота в постнатальном онтогенезе
Рецензенты:
Квочко А.Н., д.б.н., профессор, заведующий кафедрой физиологии, хирургии и акушерства ФГБОУ ВПО «Ставропольский государственный аграрный университет», г. Ставрополь;
Криворучко А.Ю., д.б.н., генеральный директор ООО НПО «Региональный центр Ветеринарной медицины», г. Ставрополь.