



Известно, что биохимический состав кормовых растений насекомых-фитофагов оказывает значительное влияние на степень предпочтения субстрата, уровень потребления кормового субстрата насекомыми, их выживаемость и степень дефолиации крон древесных растений [5]. Вследствие этого параметры энтоморезистентности растений являются ключевыми, детерминирующими как возможность реализации вспышек массового размножения лесных насекомых-фитофагов, так и степень дефолиации крон и площади очагов [2]. Понятие «энтоморезистентность» рядом авторов дифференцировано на конститутивную и индуцированную резистентность растений к насекомым [6; 10; 14]. Конститутивная энтоморезистентность (природная) является генетически детерминируемой неповреждаемостью их насекомыми. Биохимически она обусловлена содержанием в хвое и листьях первичных и вторичных метаболитов, которые обеспечивают избегание и непредпочитаемость насекомыми этих растений [10; 12]. Наиболее явным примером является содержание ряда алкалоидов в кормовом субстрате, которые токсичны для потребителей этого корма [7]. Другим важным компонентом конститутивной резистентности служит физическая защита листьев и хвои (наличие шипиков на поверхности растений, трихом, эпикутикулярного воска, жесткость листовой пластинки [6; 16; 21]. Индуцированная энтоморезистентность активируется только воздействием на растения различных стрессовых факторов (дефолиация филлофагами, инфицирование болезнями, воздействие весенне-летних засух) [10; 12]. Биохимические аспекты механизмов индуцированной энтоморезистентности в последнее время изучаются очень интенсивно [1; 3; 4; 9; 10; 14; 15; 19]. Биохимические аспекты конститутивной энтоморезистентности изучены значительно меньше. Исходя из этого биохимические аспекты конститутивной энтоморезистентности и были задачей данного исследования.
Материалы и методы
Для хроматографического анализа проводился сбор листьев березы повислой из очагов массового размножения непарного шелкопряда на пробных площадях с ежегодно регистрируемым процентом дефолиации крон и за границей очагов, где отсутствовала дефолиация. Взятие каждой пробы осуществлялось от нескольких деревьев. Затем пробы смешивались для получения средней пробы. Район очагов массового размножения находится в Каменск-Уральском районе Свердловской области. Пробные площади располагались в березняках вблизи пос. Покровское. Контрольные пробы взяты в березняках вблизи пос. Храмцовское, растущих в сходных лесорастительных условиях. В этих насаждениях периодические вспышки массового размножения непарного шелкопряда отсутствовали, заметной дефолиации не наблюдалось.
Сразу после сбора листья березы высушивали при 60 °С, затем размалывали. После этого навеску с 2 г размолотых листьев смешивали с 20 мл 95%-ного этанола. Экстракцию фенольных соединений из листьев березы проводили в обратном холодильнике на водяной бане в течение 30 мин при кипении раствора. После этого суспензию центрифугировали при 10000 g в течение 10 мин. Супернатант фильтровали через фильтр с диаметром пор 0,2 мкм. Хроматографический анализ проводили на жидкостном хроматографе Shimadzu LC-20 со спектрофотометрическим УФ-детектором. Детектирование элюента осуществляли одновременно на двух полосах поглощения: 254 и 360 нм на хроматографической колонке PerfectSil Target ODS-35мкм с обращенной фазой, размером 250 х 4,6 мм.
Градиентное элюирование проводилось в диапазоне 10-50% со скоростью 1 мл в мин при температуре 40 °С. Элюент А - ацетонитрил - 0,05 М фосфатный буферный раствор (рН=3,0); элюент В - ацетонитрил-вода (9:1). Продолжительность хроматографического анализа - 45 мин при 40 °С. Из них от 0 до 30 мин проводилось градиентное элюирование в диапазоне 10-50%, затем в течение 15 мин при концентрации 50%. Для идентификации фенольных соединений использовали вещества-свидетели фирм Fluka, Sigma, Aldrich: аскорбиновая к-та, арбутин, бензойная к-та, гиперозид, галловая к-та, гидрохинон, кверцетин, изокверцетин, кафтаровая к-та, рутин, кофейная к-та, феруловая к-та, 3-кофеоилхинная к-та, 4-кофеоилхинная к-та, 5-кофеоилхинная к-та, кемпферол, мирицетин, салицин.
Результаты
Градиентный хроматографический анализ (ВЭЖХ) образцов из листьев березы повислой, взятых как в очагах массового размножения непарного шелкопряда, так и за их границами, позволил выявить 48-54 фенольных соединения (рис. 1). Из общего количества фракций нами идентифицировано 12 фенольных соединений. Список идентифицированных нами фенольных соединений в листьях березы представлен в табл. 2. По химическому составу это были флавоноиды, фенолгликозиды, фенолкарбоновые и гидроксикоричные кислоты.
Для анализа основных, доминирующих тенденций в содержании фенольных соединений в березовых насаждениях за границами очагов нами проведен как тотальный сравнительный количественный попарный анализ основных выявленных при хроматографии пиков (за исключением минорных фракций), так и сравнительный анализ идентифицированных нами фенольных соединений (табл. 1, 2).
Тотальный попарный сравнительный анализ образцов листьев из очагов и контрольных древостоев (за границами очагов) (пиков с идентичным ВУ и спектральным соотношением (S360 /S254)) убедительно продемонстрировал, что эти образцы значительно отличались между собой (табл. 1).
Как показали результаты сравнительного изучения хроматограмм, в образцах листьев березы повислой, собранных в насаждениях за границами очагов, доминирующей тенденцией было заметное возрастание содержания фенолсодержащих соединений. Так, у 38,88% фракций их содержание заметно увеличивалось, у 33,33% их количественный состав не изменялся по сравнению с образцами из очагов массового размножения непарного шелкопряда, у 27,8% их содержание снижалось (табл. 1). При этом обращает внимание значительный уровень изменения содержания некоторых фракций (табл. 1). Для более глубокого анализа биохимических механизмов, происходящих в клетках листьев березы в насаждениях за границами очагов (с высокой природной (конститутивной) энтоморезистентностью), было целесообразно проанализировать тенденции реакций, идентифицированных нами на хроматограммах фенольных соединений, так как функции ряда из них хорошо известны.
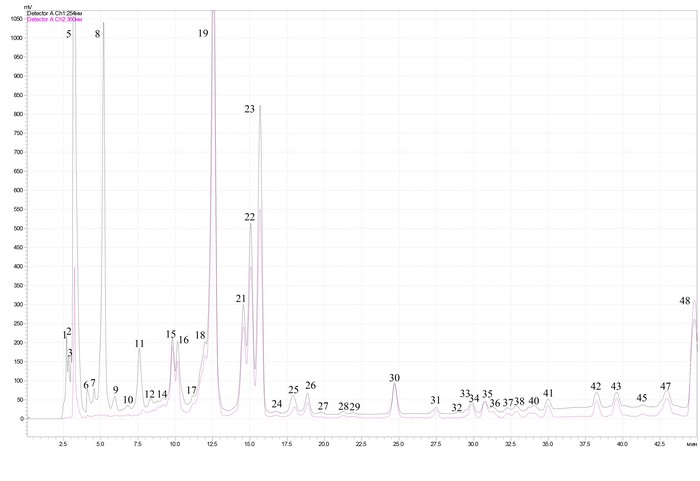
Рис. 1. ВЭЖХ фенольных соединений из листьев березы повислой в затухших очагах массового размножения непарного шелкопряда в условиях антропогенного воздействия: 1 - аскорбиновая кислота; 3 - арбутин; 5 - галловая кислота; 6 - салицин; 9 - гидрохинон; 11 - 4 - кофеоилхинная кислота; 15 - гиперозид; 16 - рутин; 22 - изокверцетин; 27 - мирицетин; 30 - кверцетин; 34 - кемпферол.
Таблица 1
Сравнительный анализ содержания фенольных соединений в листьях березы повислой в очагах массового размножения непарного шелкопряда и за их границами
|
№ п/п |
Очаг (пос. Покровское) |
Вне очага (пос. Храмцовское) |
|||
|
Абсорбция, mv (M±SD) |
Абсорбция, mv (M±SD) |
Изменение содержания фенольных соединений (в %) |
Спектральное отношение S360 /S254 |
ВУ |
|
|
1 |
259,39±26,34 |
335,16±38,06 |
+29,21 |
0,008 |
2.6 |
|
2 |
113,46±15,59 |
186,81±27,09 |
+64,65 |
0,038 |
2,82 |
|
3 |
284,33±32,65 |
374,72±27,40 |
+31,79 |
0,07 |
3,0 |
|
5 |
4100,0±109,54 |
4625,0±236,90 |
+12,81 |
0,61 |
3,5 |
|
6 |
780,50±41,50 |
926,1±43,29 |
+18,65 |
0,02 |
3,75 |
|
7 |
114,27±14,67 |
137,98±12,21 |
+20,78 |
0,001 |
4,3 |
|
8 |
497,08±27,34 |
478,19±54,91 |
--- |
0,14 |
4,8 |
|
9 |
210,65±17,33 |
145,05±15,27 |
--- |
0,004 |
5,5 |
|
10 |
465,66±60,208 |
296,43±101,33 |
--- |
0,81 |
6,3 |
|
11 |
460,71±25,56 |
380,82±47,52 |
-17,34 |
0,30 |
6,7 |
|
12 |
4300,0±246,21 |
1517,85±161,96 |
-64,70 |
0,64 |
7,5 |
|
13 |
169,62±49,98 |
153,88±25,72 |
--- |
0,57 |
7,8 |
|
14 |
296,51±19,32 |
303,55±39,54 |
--- |
0,29 |
9,4 |
|
15 |
262,36±18,52 |
170,26±17,63 |
--- |
0,98 |
10,5 |
|
16 |
60,46±6,83 |
66,40±4,76 |
+9,82 |
0,36 |
10,95 |
|
17 |
128,98±12,82 |
136,52±8,85 |
--- |
0,37 |
11,3 |
|
18 |
84,32±6,28 |
217,50±18,75 |
+157,95 |
0,88 |
14,5 |
|
19 |
406,97±32,55 |
746,97±81,12 |
+83,54 |
0,87 |
14,8 |
|
20 |
6510,40±319,08 |
5960,42±482,12 |
--- |
0,28 |
15,35 |
|
21 |
607,14±75,23 |
490,36±25,56 |
-19,20 |
0,46 |
16,5 |
|
22 |
1103,02±142,95 |
545,33±39,56 |
-50,56 |
0,45 |
18,5 |
|
23 |
2048,07±255,93 |
1034,28±115,80 |
-49,49 |
0,43 |
18,69 |
|
24 |
34,70±4,18 |
20,67±2,82 |
-40,43 |
1,04 |
19,8 |
|
25 |
30,49±12,34 |
40,46±4,72 |
+32,70 |
0,48 |
19,85 |
|
26 |
40,66±6,45 |
71,97±6,46 |
+77,0 |
0,67 |
20,0 |
|
27 |
65,66±7,29 |
83,79±4,41 |
+27,61 |
0,67 |
20,5 |
|
28 |
94,50±7,82 |
65,66±4,29 |
-30,52 |
0,63 |
21,6 |
|
29 |
143,82±20,61 |
142,48±6,44 |
--- |
0,97 |
25,5 |
|
30 |
38,45±4,17 |
33,218±6,81 |
--- |
0,65 |
30,2 |
|
31 |
82,14±7,26 |
129,12±13,49 |
+57,20 |
1,22 |
32,5 |
|
32 |
78,29±10,53 |
112,64±8,37 |
+43,88 |
0,62 |
33,6 |
|
33 |
80,22±12,65 |
74,17±5,04 |
--- |
0,68 |
39,0 |
|
34 |
97,53±11,55 |
44,78±5,11 |
-54,09 |
0,53 |
40,6 |
|
35 |
60,99±4,50 |
44,50±2,40 |
-27,04 |
0,60 |
42,2 |
|
36 |
72,81±5,51 |
66,48±6,63 |
--- |
0,51 |
42,7 |
|
37 |
661,50±42,17 |
345,05±28,86 |
-47,84 |
0,88 |
45,0 |
Очевидно, что наибольший интерес представляла группа идентифицированных нами фенольных соединений, содержание которых было выше, чем в потенциально низкорезистентных насаждениях (в очагах).
Таблица 2
Сравнительный анализ содержания идентифицированных фенольных соединений в листьях березы повислой в очагах массового размножения непарного шелкопряда и за их границами
|
№ п/п |
Очаг (пос. Покровское) |
Вне очага (пос. Храмцовское) |
|||
|
|
Абсорбция, mv (M±SD) |
Абсорбция, mv (M±SD) |
Изменение содержания фенольных соединений (в %) |
Спектральное отношение S360 /S254 |
ВУ |
|
Аскорбиновая к-та |
259,39±26,34 |
335,16±38,06 |
+29,21 |
0,008 |
2.6 |
|
Арбутин |
284,33±32,65 |
374,72±27,40 |
+31,79 |
0,07 |
3,08 |
|
Галловая к-та |
780,50±41,50 |
926,1±43,29 |
+18,65 |
0,02 |
3,62 |
|
Салицин |
114,27±14,67 |
137,98±12,21 |
+20,78 |
0,001 |
4,3 |
|
Гидрохинон |
210,65±17,33 |
145,05±15,27 |
--- |
0,004 |
5,5 |
|
4-кофеоилхинная кислота |
460,71±25,56 |
380,82±47,52 |
-17,34 |
0,30 |
6,7 |
|
Рутин |
262,36±18,52 |
170,26±17,63 |
--- |
0,98 |
10,5 |
|
Гиперозид |
84,32±6,28 |
217,50±18,75 |
+157,95 |
0,88 |
14,5 |
|
Изокверцетин |
406,97±32,55 |
746,97±81,12 |
+83,54 |
0,87 |
14,8 |
|
Мирицетин |
34,70±4,18 |
20,67±2,82 |
-40,43 |
1,04 |
19,8 |
|
Кверцетин |
143,82±20,61 |
142,48±6,44 |
--- |
0,97 |
25,4 |
|
Кемпферол |
82,14±7,26 |
129,12±13,49 |
+57,20 |
1,22 |
32,5 |
Как показали результаты сравнительного анализа, содержание аскорбиновой кислоты в пробах возрастало на 29,21%, арбутина - на 31,79%, галловой кислоты - на 18,65%, салицина - на 20,78%, гиперозида - на 157,95%, изокверцетина - на 83,54%, кемпферола - на 57,2% (табл. 2). В их состав входили флавоноиды (3), фенолгликозиды (2), фенолкарбоновые кислоты (1), витамины (1) (табл. 2).
Известно, что эффективными антифидантными свойствами из этой группы фенольных соединений, в первую очередь, обладают алкалоиды и фенолгликозиды [7; 11]. Поэтому повышение уровня содержания таких фенолгликозидов, как арбутин и салицин, можно однозначно рассматривать как аргумент в пользу наличия более высокого уровня природной (конститутивной) энтоморезистентности в насаждениях за границами очагов. Следует также учитывать, что в течение весенне-летнего сезона 2014 г. в Каменск-Уральском районе Свердловской области засухи отсутствовали. Следовательно, отсутствовала и реакция древостоев на фактор абиотического стресса (засуху). Фактор дефолиации как в контрольных насаждениях, так и в насаждениях в затухших очагах, также, полностью отсутствовал.
Содержание ряда других фенольных соединений в листьях, наоборот, заметно снижалось. Это 4-кофеоилхинная кислота (на 17,34%) и мирицетин (на 40,43%). Ряд авторов предполагает, что мирицетин из флавоноидов обладает наиболее активным антиоксидантным эффектом [19]. Поэтому его снижение можно рассматривать как следствие отсутствия активных процессов оксидативного стресса в насаждениях за границами очагов.
Содержание ряда других фенольных соединений в образцах из березовых насаждений за границами очагов не изменялось. Это гидрохинон, рутин, кверцетин (табл. 2).
Ранее нами было показано, что в этих насаждениях были обнаружены заметные различия в уровне реакции на фактор абиотического стресса (весенне-летнюю засуху) [4]. Древостои в затухших очагах отличались от контрольных (за границами очагов) тем, что их уровень реакции на этот стресс-фактор был значительно выше. В результате хроматографический спектр экстракта из листьев количественно значительно отличался от контроля [4].
Как показали результаты, из общего количества фенольных соединений, содержание которых в контроле возрастало, преобладали флавоноиды и фенолгликозиды. Известно, что одной из основных функций флавоноидов в клетках растений является антиоксидантная [13; 17; 20]. Кроме того, некоторые авторы показали, что, например, листья дуба у деревьев с высокой энтоморезистентностью отличались повышенным содержанием полифенолов, флавоноидов, танина и лигнина [18]. Полученные нами результаты показали, что из всех групп фенольных соединений, содержание которых возрастало в листьях насаждений за границами очагов, содержание таких флавоноидов, как гиперозид, изокверцетин и кемпферол, возрастало в наибольшей степени (от 57 до 157%). Для сравнения сопоставим особенности реакции этих соединений на фактор дефолиации и на стресс (весенне-летнюю засуху). Как показали результаты, полученные нами ранее, проявлением индуцированной фактором дефолиации крон резистентности было заметное снижение содержания одних фенольных соединений, по сравнению с контролем и возрастанием других [3]. Так, в результате дефолиации и активизации функции замедленной индуцированной резистентности в листьях березы в очагах значительно возрастало содержание гиперозида, рутина, галловой, кофейной, аскорбиновой и феруловой к-т, и снижалось содержание 3-КФК, 4-КФК и 5-КФК [3]. Дополнительно проведенный анализ содержания в пробах таких фенолгликозидов, как арбутин и салицин, показал, что содержание арбутина в листьях образцов из очагов составляло 503,66±18,52 mv, тогда как в листьях за границами очагов - 328,34±25,57 mv (на 53,39% выше). Содержание салицина соответственно 285,47±19,47 mv и 179,08±9,71 mv (на 59.41% выше). Это подтверждает, что в результате активации замедленной индуцированной резистентности в листьях значительно возрастает содержание ряда фенолгликозидов, обладающих антифидантной активностью. Ранее нами показано, что в результате воздействия фактора абиотического стресса (засухи) в насаждениях в затухших очагах заметно снижалось содержание аскорбиновой, галловой к-т, а также кверцетина, изокверцетина, мирицетина [4]. Возможно, что это оказывает влияние на улучшение кормовой ценности субстрата. Но учитывая, что многие флавоноиды являются мощными антиоксидантами, нельзя исключить также и то, что частично изменение активности их синтеза обусловлено и реализацией антиоксидантной функции в условиях воздействия как фактора абиотического стресса (засухи), так и дефолиации. Известно, что вследствие воздействия этих факторов возникает повышенная оксидативная активность в клетках листьев, вследствие чего активизируется антиоксидантная активность клеток, в том числе и за счет возрастания активности фенолсодержащих соединений, одной из важных функций которых является антиоксидантная. Это существенно затрудняет возможность интерпретации полученных результатов.
Синтез ряда других фенольных соединений, наоборот, возрастает в насаждениях в затухших очагах. Это гиперозид, рутин, кофейная к-та, гидрохинон, 4-КФК и 5-КФК. Он также может быть обусловлен реализацией антиоксидантной функции. Обращает внимание частичное сходство состава фенольных соединений, содержание которых заметно возрастает в результате активации фактора индуцированной дефолиацией резистентности и, предположительно, параметров конститутивной энтоморезистентности в насаждениях вне очагов.
Таким образом, можно предполагать, что отдельные фенольные соединения могут участвовать и в реализации индуцированной резистентности, и являться компонентом конститутивной (природной) энтоморезистентности. Подобное предположение уже высказывалось ранее другими исследователями [8].
Учитывая то, что в составе фенольных соединений в листьях березы за границами очагов доминируют соединения с заметным увеличением их содержания по сравнению с насаждениями в границах очагов, а также то, что в их состав входят идентифицированные фенолгликозиды с антифидантными свойствами, можно предполагать, что они могут быть показателями высокого уровня конститутивной энтоморезистентности насаждений.
Проект выполнен при финансовой поддержке РФФИ (грант № 13-04-96063) и правительства Свердловской области.
Рецензенты:
Усольцев В.А., д.с.-х.н., профессор, профессор Уральского государственного лесотехнического университета, г. Екатеринбург.
Петрова И.В., д.б.н., заведующий лабораторией популяционной биологии растений и динамики леса ФГБУН «Ботанический сад Российской академии наук», г. Екатеринбург.