



Введение
Длиннохвостая синица на территории Чувашской Республики представлена подвидом Aegithalos caudatus caudatus и является широко распространенным видом в лесных массивах, как исследуемого региона, так и европейской части России, который встречается здесь в течение всего года. Несмотря на то, что на территории России этот вид является обычным, экология вида на данной территории изучена очень слабо. Наиболее подробные популяционные исследования выполнены на подвиде A. c. rosaceus в Англии [7], в центральной части России экологические особенности длиннохвостой синицы изучены фрагментарно. Основные публикации рассматривают детально гнездовую биологию вида, достаточно подробно изучена динамика осенних миграций. Было установлено, что длиннохвостым синицам из европейских популяций свойственны резкие колебания численности в период осенних миграций, которые происходят в отдельные годы с разной периодичностью, что характеризует их как нерегулярных мигрантов [8]. В связи с этим представляется особенно актуальным рассмотрение вопроса о причинах таких резких перепадов численности вида.
Цель исследования: определить особенности ее многолетней динамики численности длиннохвостых синиц в Чувашской Республике и оценить ее стабильность.
Материалы и методы исследования
Исследования проводили с 1999 по 2010 годы на стационаре, расположенном на территории Природного парка «Заволжье» Чувашской Республики в окрестностях озера Малое Лебединое. На стационаре в полосе зарастания озера была установлена рабочая площадка площадью около 2 га, где в строго фиксированных местах располагались паутинные сети, которыми проводили отлов птиц. Отлов и кольцевание длиннохвостых синиц проводили весной (с конца апреля до середины мая) и с середины июня до конца октября – начала ноября стандартными паутинными сетями. Количество сетей в разные годы варьировало (обычно 5-10, в некоторые годы - до 20). Отметим, что для получения объективных показателей нами был установлен показатель «уловистость», который позволяет скорректировать возможные ошибки в анализе динамики отловов, связанные с тем, что в разные годы количество сетей, которыми проводился отлов птиц, был непостоянен. Показатель уловистость высчитывался по формуле:
Уловистость= ![]() х100
х100
При мечении птиц использовали стандартные алюминиевые кольца Центра кольцевания РАН и цветные пластиковые кольца. Отловленные птицы подвергались стандартной процедуре прижизненной обработки. Всего были использованы материалы по отловам и прижизненному анализу 2618 длиннохвостых синиц.
Результаты исследования и их обсуждение
По результатам наших исследований на территории Чувашской Республики у популяции длиннохвостых синиц происходят значительные колебания численности из года в год. Наибольшая численность вида была отмечена в данном регионе в 2001 году (539 особей), а в 2008 году на исследуемой территории данный вид не отмечался вовсе (рисунок 1). Показатель уловистости при этом коррелирует с количеством длиннохвостых синиц, отловленных во время послегнездового периода (r=0,77; p<0,01; n=10).
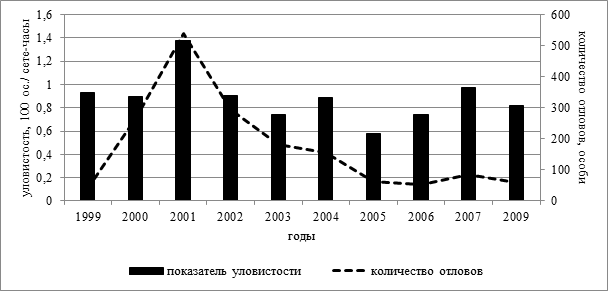
Рисунок 1. Динамика отловов длиннохвостых синиц и показатель уловистости вида в разные годы
Подобные колебания численности исследуемого вида отмечаются в работах, проведенных в других регионах Среднего Поволжья [1]. Таким образом, мы предполагаем, что обнаруженный нами всплеск численности в 2001 году имел не случайный характер, а был связан с тем, что в этот год происходили массовые осенние миграции вида на территории Среднего Поволжья, в которые были вовлечена значительная часть исследуемой нами популяции длиннохвостых синиц. Стоит отметить, что подобное явление, когда подобные массовые миграции затрагивали сразу несколько прилегающих территорий, уже отмечалось ранее в литературе. Так, в 1959 году, когда в Ленинградской области проходила инвазия вида по всему региону, аналогичные массовые всплески мигрирующих птиц были зафиксированы в Вологодской и Псковской областях, в Республике Карелия и ряде других районов [3].
Тем не менее, в более отдаленных от исследуемого нами региона областях, колебания численности длиннохвостых синиц носили отличный, но при этом схожий характер: на Ладожской орнитологической станции наибольшая численность ополовников была зафиксирована в 2000-01 годах [2], на Куршской косе всплеск отловов был отмечен в 2000 году [6]. Мы считаем, что отмеченная другими исследователями инвазия вида, происходившая в северо-западном регионе в 2000 году, затронула частично и популяцию ополовников Среднего Поволжья. Это вынудило часть особей популяции Чувашской Республики покинуть места обитания и начать перемещения на значительные расстояния в западном направлении. Именно в тот год, молодая длиннохвостая синица, окольцованная на нашем стационаре 25 июня 2000 г., была обнаружена по данным Центра кольцевания РАН спустя некоторое время - 7 октября 2000 года на станции кольцевания Ventas Ragas (Латвия) [9].
Таким образом, увеличение численности и массовые перемещения ополовников в разных частях ареала в различные годы в Европейской России происходят не синхронно, а независимо друг от друга. Однако массовые миграции вида, проходящие в отдельном регионе, могут затрагивать обширную площадь, вынуждая тем самым птиц из соседних территорий совершать значительные перемещения в едином для всей миграции направлении. В подобных случаях отдельные птицы из популяции Чувашской Республики оказываются вовлеченными в массовые миграции, происходящие в более отдаленных регионах.
Мы также провели анализ стабильности численности исследуемого вида в разные годы. Количественную оценку стабильности численности можно дать, используя статистические показатели вариации численности – разницу между ее максимальными и минимальными значениями (размах колебаний), оценку среднеквадратического отклонения, коэффициент вариации и некоторые другие. В нашем случае для этой цели мы использовали относительную величину – индекс стабильности численности IS, который рассчитывается по следующей формуле [4]:
![]() ;
;
![]() , если
, если ![]() ≤
≤![]() ,
,
![]() , если
, если ![]() ≥
≥ ![]() ,
,
![]() – i-e значение оценки численности (плотности);
– i-e значение оценки численности (плотности); ![]() – следующее после i-го сопряженное во времени значение оценки численности (плотности); N – количество вариантов оценок численности; M – количество разобщенных некратным временным промежутком массивов вариантов оценок численности (плотности); если массив один, не разобщен, то m=1.
– следующее после i-го сопряженное во времени значение оценки численности (плотности); N – количество вариантов оценок численности; M – количество разобщенных некратным временным промежутком массивов вариантов оценок численности (плотности); если массив один, не разобщен, то m=1.
В каждом массиве должно быть вариантов больше одного. Если ![]() =
= ![]() = 0, то m увеличивается на количество таких случаев, а соответствующее
= 0, то m увеличивается на количество таких случаев, а соответствующее ![]() в расчете не участвует.
в расчете не участвует.
Ошибка индекса (SE) рассчитывается по формуле:
![]() ,
,
где
![]() =
= ![]() ,
,
если n – m > 30, и
![]() =
= ![]() ,
,
если n – m < 30.
Индекс стабильности численности (плотности) изменяется в пределах от 0 до 1 (0 ≤ IS ≤ 1). Если IS = 0, то население вида данной территории крайне нестабильно; если IS = 1, то население максимально стабильно и не изменяется во времени.
Полученные данные стабильности численности отражены на рисунке 2. Видно, что этот показатель варьировал в разные годы – наименьшая стабильность численности длиннохвостых синиц в Чувашской Республике была в 2002 и 2007 году, наиболее стабильная численность вида отмечалась в 2003 году.
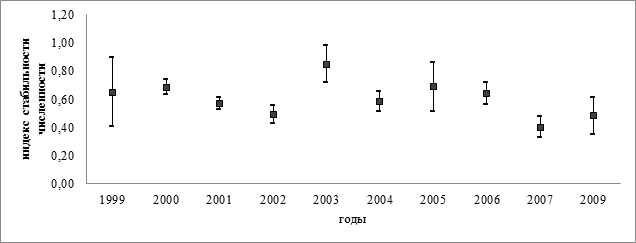
Рисунок 2. Динамика индекса стабильности численности длиннохвостых синиц по результатам отловов в разные годы (IS ± SE)
Подобные колебания стабильности численности могут быть следствием различных процессов, происходящих в популяции, однако выявить связь стабильности численности с динамикой отловов не удалось (r=-0,15; p>0,05; n=10).
Мы решили провести анализ, может ли быть динамика стабильности численности обусловлена температурными показателями в исследуемом регионе, поскольку известно, что температурный режим может влиять на миграционную активность птиц и приводить к значительному колебанию их численности (Соколов, 2007). Для этого мы сравнили полученные результаты динамики индекса стабильности численности с различными показателями температурного режима в Чувашской Республике. В результате были выявлены достоверные зависимости стабильности численности от средних минимальных температур летних месяцев и средних максимальных температур воздуха осенью (рисунки 3 и 4). В те годы, когда стабильности численности была наибольшей, отмечались более низкие температуры в летние месяцы, но были более высокие температуры в осенние месяцы.
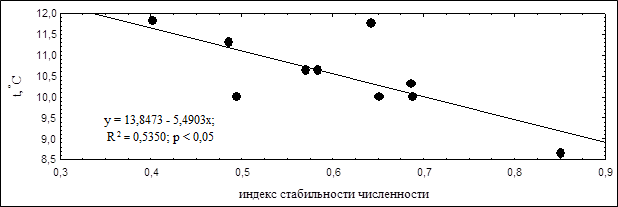
Рисунок 3. Корреляция средних минимальных температур летних месяцев с индексом стабильности численности
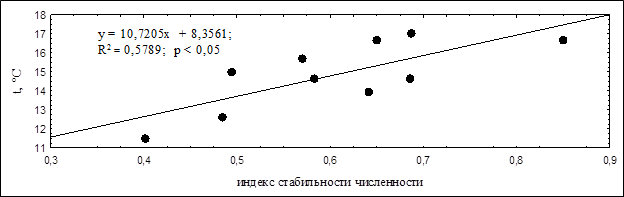
Рисунок 4. Корреляция средних максимальных температур осенних месяцев с индексом стабильности численности
Вероятно, обратная корреляция индекса стабильности численности от минимальных летних температур может быть связана с тем, что в годы, когда температура воздуха не снижается значительно и сохраняет высокие показатели, птицам становится сложнее поддерживать уровень теплообмена с окружающей средой, что может приводить к обезвоживанию тела и его перегреву. Эти факторы повышают риск гибели птиц во время летних кочевок, что, в свою очередь, отражается в снижении индекса стабильности численности.
Наоборот, выявленная нами положительная связь индекса стабильности численности с максимальными температурами в осенние месяцы может быть объяснена следующими причинами: в годы, когда максимальные температуры воздуха выше, во-первых, проще найти себе пищу (у насекомых продлевается период активности), а во-вторых, уменьшаются затраты на поддержание температуры тела и снижается риск гибели из-за переохлаждения.
Отметим, что ранее уже было показано наличие связей между интенсивности осенней миграции вида и температурными показателями. В исследованиях, проведенных на Куршской косе, была установлена значимая положительная связь между осенней численностью птиц и весенними температурами воздуха для большей части Европейской части России [5]. Для ополовников, обитающих на территории соседней Татарской Республики, была установлена корреляция численности вида с температурными показателями прошлогоднего зимнего периода (ноябрь - март) [1]. Результаты нашей работы позволяют говорить не только о том, что динамика осенних миграций, но в целом стабильность численности вида подвержена влиянию температурного режима летних и осенних месяцев.
Выводы
В результате нашей работы мы установили, что численность длиннохвостых синиц по результатам отловов в разные годы подвержена значительным колебаниям. Наибольшая численность вида в Чувашской Республике была отмечена в 2001 годы, в 2008 году птицы не отмечались вообще. В отдельные годы наблюдаются резкие всплески численности вида, которые затрагивают соседние области. При этом массовые осенние миграции вида, происходящие в отдалённых регионах, затрагивают часть птиц из местной популяции.
Стабильность численности ополовников подвержена влиянию температурного режима в регионе, она зависит от минимальных среднемесячных показателей в летние месяцы и от максимальных среднемесячных показателей осенью.
Рецензенты:
Белова Н.А., д.б.н., ведущий научный сотрудник, Федеральное государственное бюджетное учреждение науки «Институт теоретической и экспериментальной биофизики РАН» (ИТЭБ РАН), г.Пущино.
Гудков С.В., д.б.н., ведущий научный сотрудник, Федеральное государственное бюджетное учреждение науки «Институт теоретической и экспериментальной биофизики РАН» (ИТЭБ РАН), г.Пущино.